






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
缺硒条件下诱导Nrf2靶基因
发表于:2021-09-01 作者:admin 来源:本站 点击量:23146
原文:M Müller, Banning A , R Brigelius-Flohé, et al. Nrf2 target genes are induced under marginal selenium-deficiency[J]. Genes & Nutrition, 2010, 5(4):297-307.
翻译:
关键词:硒;Nrf2; II相酶; 肠;氧化应激
引言
在哺乳动物中,必需的微量元素硒作为硒代半胱氨酸并入硒蛋白活性中心发挥作用。人类有25个基因编码硒蛋白[30],其中许多涉及氧化还原过程[35],包括谷胱甘肽过氧化物酶(GPx)[8]和硫氧还蛋白还原酶(TrxR)[14]。欧洲人口中存在的少量硒缺乏症可能伴随着轻微的氧化状态[45]。 硒蛋白的表达取决于细胞中硒的状态。但是,一些微阵列研究表明,硒的可用性也会影响非硒蛋白的表达(在[13]中进行了综述)。仍然缺乏基本机制。硒状态码经常影响抗氧化剂防御酶和II期系统的酶[13,44]。一方面,大剂量的硒刺激诸如谷胱甘肽S转移酶(GST)和NADPH:醌氧化还原酶(NQO1)等II期酶的活性。这种作用可以分别通过硒代半胱氨酸硒结合物[53]和二甲基二硒化物[58]来介导。另一方面,有强有力的证据证明在硒缺乏时会诱导抗氧化剂和II期酶的产生。早在1970年代末期,据报道血红素加氧酶1(HMOX1)[12]以及GST活性在缺硒的大鼠肝脏中有所升高[32]。硒缺乏症中抗氧化剂和II期酶的补偿性上调的想法进一步得到了小鼠的发现,该小鼠具有针对器官的靶向去除编码硒代半胱氨酸tRNA(Trsp)的基因。肝脏中硒蛋白的完全丧失导致了GSTPi,NQO1和HMOX1的诱导[52]。 参与细胞抗氧化剂防御和II期排毒的大多数蛋白质基因是由氧化还原和亲电敏感的转录因子NF-E2相关因子(Nrf2)调控的(在[16]中进行了综述),该因子与顺式作用的抗氧化剂或亲电反应元件(ARE / EpRE)[48,56]。 Nrf2-/-小鼠对化学诱导的毒性和肿瘤发生敏感[43]。另外,Nrf2-/-小鼠对氧化应激高度敏感[1],并具有较低的基础谷胱甘肽(GSH)水平[47]。在不受刺激的条件下,Nrf2保留在与Keap1结合的胞质溶胶中,Keap1充当基于Cul3的E3泛素连接酶的底物衔接子,并靶向Nrf2进行降解。靶基因的反式激活是响应活性氧(ROS)或亲电试剂而引起的,这些试剂修饰Keap1的易感硫醇基团,导致核Nrf2蛋白水平迅速提高(在文献[38]中进行了综述)。 Trsp和Nrf2的同时破坏消除了II期酶的诱导,从而验证了Nrf2在响应硒蛋白丢失中的作用[52]。与饲喂对照饮食的小鼠相比,饲喂缺硒饮食的ARE报告小鼠的ARE驱动的报告基因活性显著增强[9]。另外,在缺硒的野生型小鼠中观察到的GST和NQO1活性增加在Nrf2-/-小鼠中无法检测到[9]。这提供了饮食中硒缺乏与Nrf2激活之间的第一个直接联系。 基于这些结果,当前的研究集中在边缘缺硒饮食对Nrf2调控基因表达的影响上。贫硒饮食中小鼠的建议每日允许摄入量(RDA)为一半[28],反映出生理状况可能会因改变营养习惯或季节性食物变化而同样发??生。分析的目标器官是结肠和十二指肠的肠段,这是抵抗异种生物胁迫的第一道防线,还有肝脏,肝脏高度表达II期酶。先前研究[28]中获得的微阵列数据的途径分析未表明Nrf2途径受到影响,因为所用程序GenMAPP不包含该途径的MAPP(微阵列途径概况)。因此,手动鉴定了48个Nrf2靶基因和12个经典II期酶,并通过qPCR证实了有趣的候选基因。
材料和方法
动物实验
从先前报道的动物中获取组织样品[28]。对雄性C57BL / 6 J小鼠(3-4周龄)饲喂贫硒(0.086 mg Se / kg)或通过混合硒代蛋氨酸(Acros,比利时的盖尔(Geel)陷入贫困的人(德国的拉格(Altromin))。喂养6周后,麻醉的动物因颈脱位而被处死。冷冻固定在液氮中的血浆和组织储存在-80°C下。每组12只动物的硒状态以血浆硒含量和肝GPx活性为特征,发现在中等硒缺乏时分别降低至硒充足的13%和35%[28]。
RNA分离
在液氮冷却下将组织研磨。将20–30 mg的粉末悬浮在800μl的冷Trizol(Invitrogen,卡尔斯鲁厄,德国)中,并用组织裂解器(Qiagen,Hilden,德国)以30 Hz匀化2×2分钟。根据制造商的说明,使用Trizol方案分离RNA。用10 U RQ1 DNase(Promega,曼海姆,德国)消化基因组DNA,并通过苯酚-氯仿提取纯化RNA。使用NanoDrop ND-1000(Peqlab Biotechnologie GmbH,德国埃尔兰根)测量RNA浓度。
芯片分析
如前所述[28],使用Mouse 44 K芯片(Agilent Technologies,Böblingen,德国)进行芯片分析。Student’s t-test检验可鉴定出显著调控的基因(P <0.05)。 实时定量PCR 用150 fmol oligo(dT)15引物和180 U莫洛尼鼠白血病病毒逆转录酶(Promega)反转录RNA(3μg),总体积为45μl。
实时PCR
(Mx3005PTM QPCR系统,Stratagene,阿姆斯特丹,荷兰)使用SYBR Green I(Molecular Probes,Eugene,USA)作为荧光报告物,在25μl反应混合物中以1μl十倍稀释的cDNA一式三份进行。所有PCR反应的退火温度分别为60和62°C(表1)。 PCR产物的定量范围为1×103至1×109个拷贝的标准曲线。通过将至少一种引物置于PerlPrimer v1.1.14的外显子/内含子边界上,设计引物(表1,Sigma–Aldrich,Taufkirchen,德国)对cDNA具有特异性。参考基因Rpl13a和Hprt1的平均值用于在结肠和肝脏中qPCR结果的标准化,而Rpl13a仅用于十二指肠。
表1. 所用底物序列表
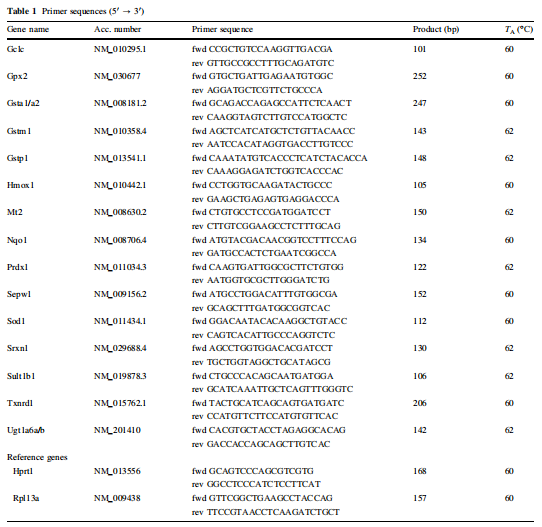
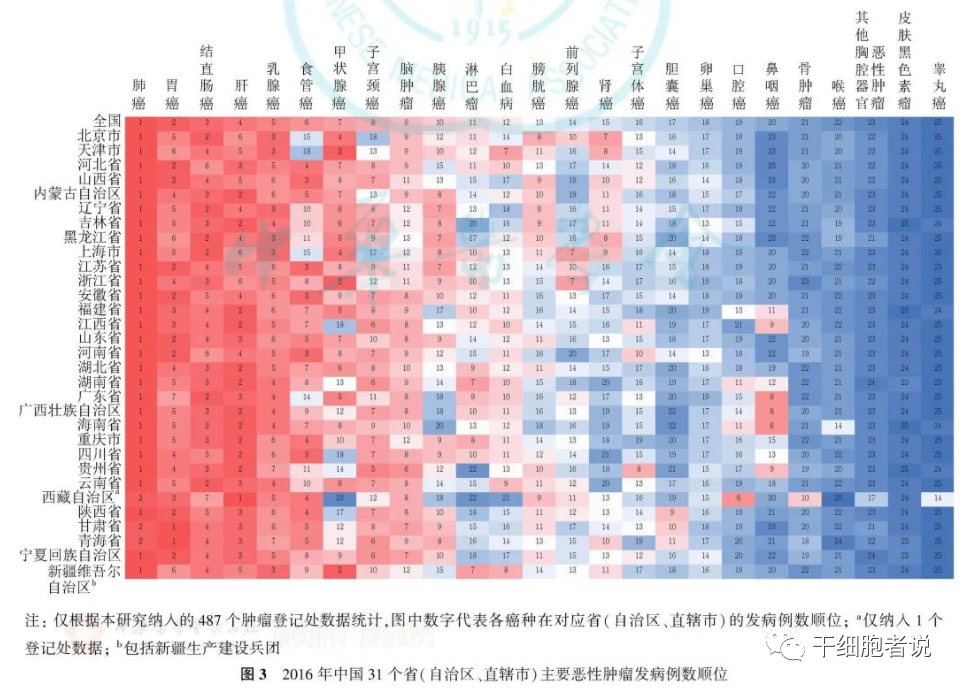
表2. 微阵列分析测量硒对结肠Nrf2靶基因作用


组织裂解液的制备
将10 mg磨碎的组织悬浮于含有2μl蛋白酶抑制剂混合物III(Calbiochem,Bad Soden,Germany)的250μl均质缓冲液(100 mM Tris-HCl,300 mM KCl,0.01%Triton X-100)中并用组织裂解器(Qiagen)在30 Hz下持续2×2分钟。裂解液在21,000×g和4°C下离心15分钟,然后根据Bradford [7]评估蛋白质含量。
蛋白质印迹
将等分试样(20μg蛋白/泳道)进行SDS-PAGE,并按[5]所述进行印迹。用兔抗人NQO1抗血清(1:3,000; ab34173,Abcam,剑桥,英国)检测到NQO1。通过兔抗人β-肌动蛋白抗血清(1:5,000; ab8229,Abcam)检测到的β-肌动蛋白用作内部对照。使用过氧化物酶偶联的抗兔抗体(1:50,000; Chemicon,霍夫海姆,德国)作为二抗。检测是在以SuperSignal West Dura(德国波恩的Perbio)为底物的Fuji LAS3000-CCD系统中完成的。用Aida / 2D光密度测定法4.0软件(Raytest,Straubenhardt,德国)对条带进行定量。
NQO1活动
根据[42]测量双香豆酚敏感的NQO1活性。在590 nm的微量滴定板吸光度读取器上追踪由甲萘醌介导的3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(MTT)的还原(Synergy 2,Biotek Instruments GmbH,Bad Friedrichshall,德国),将不同的填充量校正为1厘米的路径长度。将3μl裂解物与190μl反应缓冲液(25 mM Tris-HCl,pH 7.4,0.665 mg / l BSA,0.01%Tween 20,5μMFAD,1 mM葡萄糖-6-磷酸盐,30μM一起用于测定NADP,0.72 mM MTT,0.3 U / ml 6磷酸葡萄糖脱氢酶,50μM甲萘醌),总体积为250μl。 NQO1的活性计算为有和没有双香豆酚时MTT还原速率之间的差异,使用的还原MTT的消光系数在590 nm处估计为11,961(mol / l)-1×cm-1。
统计分析
比较两组,通过未配对的Student’s t-test(GraphPadPrism®5.0版,美国加利福尼亚州圣地亚哥)计算出显著差异。 P值<0.05被认为具有统计学意义。
结果
两个Nrf2硒蛋白靶基因GPx2和TrxR1是特殊情况,因为它们的翻译依赖于硒的可用性及其在层次结构中的排名。 高硒蛋白GPx2 [57]和TrxR1 [10]的mRNA在缺硒结肠中显著降低,GPx2受影响小于TrxR1。 相反,两种酶的mRNA在硒缺乏十二指肠中增强(图3a,b)。 硒缺乏时GPx2 mRNA的上调已经在细胞培养中显示[57]。 在限制硒条件下mRNA的稳定性使得在补充硒后能够在缺硒细胞中快速重新合成GPx2 [57]。 两种观察都可以作为硒蛋白质层次中GPx2高级别的证据。NQO1 RNA在缺硒十二指肠中的上调与NQO1蛋白表达和活性相关
在蛋白质和活性水平上进一步分析NQO1作为Nrf2靶基因的原型。 在十二指肠中,NQO1蛋白和活性在缺硒组中显著增强(图4),与增强的mRNA相关(图2a)。 在结肠中,蛋白质水平和活性都没有改变,表明NQO1 mRNA的轻微增加不会通过增加的翻译和活性反映出来。
讨论
本研究是动物实验的一部分,其中给小鼠喂食符合RDA的小鼠或硒含量降低至约50%的饮食[28]。虽然硒摄入量仅适度减少,但四种硒蛋白SelW,GPx1,SelH和SelM的RNA表达明显下调,Wnt途径的基因在结肠中上调[28]。对微阵列数据的人工分析表明,Nrf2靶基因和II期酶也受硒状态的调节;在结肠中表达的60个基因中,41个被上调占68%(表2)。通过qPCR证实了12种用于抗氧化防御和II期酶的所选基因的表达(图1和2)。 Nrf2靶基因在边缘硒缺乏中的一致上调是高度相关的,因为次优的硒供应也可能是由人类经常消耗的低硒饮食造成的[45]。边缘硒缺乏的建立与大多数其他研究高硒补充剂或严重缺硒的研究相反。
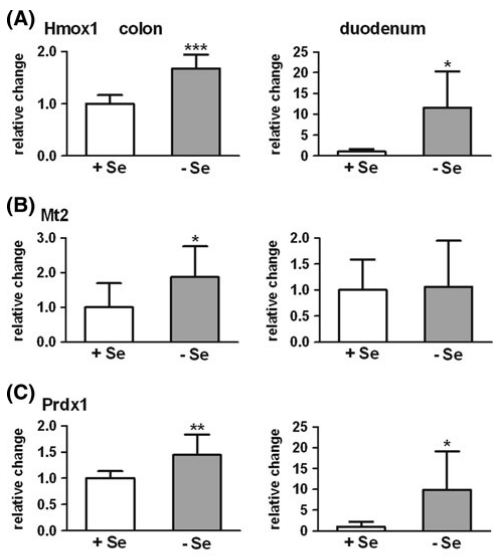

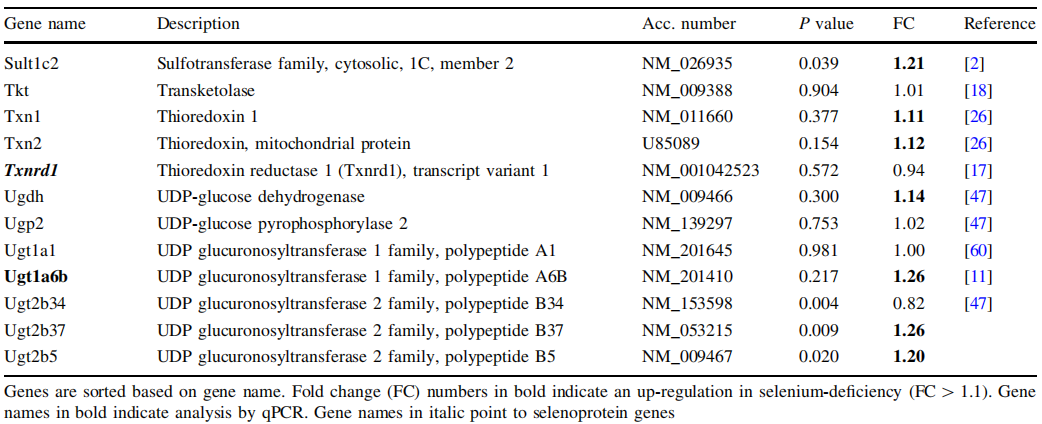
图1.硒缺乏(-Se)小鼠与相对于不缺硒(+Se)小鼠的结肠(n = 9)和 十二指肠(n = 6)中抗氧化酶的mRNA表达分析。 a血红素加氧酶1(Hmox1),b金属硫蛋白2(Mt2),c peroxiredoxin 1 (Prdx1), d sulfifiredoxin(Srxn1),e超氧化物歧化酶1(Sod1),f c-谷氨酰半胱氨酸合成酶,催化亚基(Gclc)。基因表达通过qPCR分析,并在十二指肠采用Rpl13a进行标准化。 在结肠中参考Hprt1和Rpl13a进行平均。 * P< 0.05; ** P< 0.01; 设置+Se为1,采用t-test分析方法进行分析
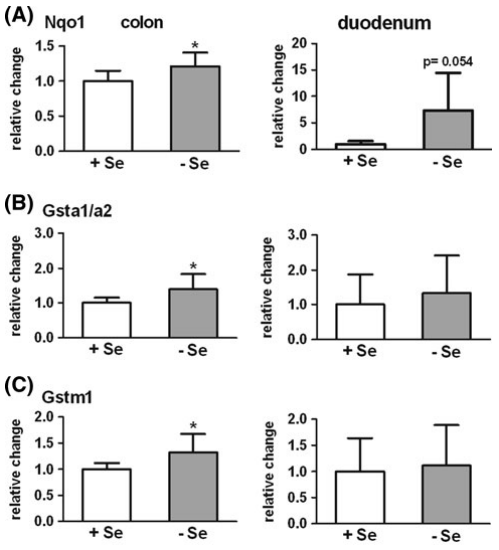
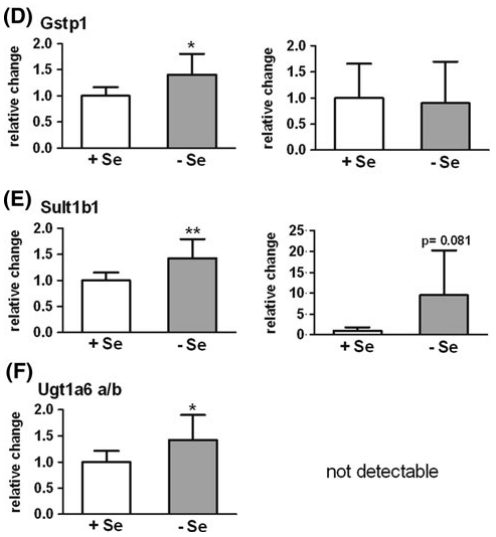
图2.硒缺乏(-Se)小鼠与相对于不缺硒(+Se)小鼠的结肠(n = 9)和 十二指肠(n = 6)中II相酶的mRNA表达。NADPH:醌氧化还原酶(Nqo1), b谷胱甘肽S转移酶a1 / a2(Gsta1 / a2),c谷胱甘肽S转移酶m1(Gstm1),d谷胱甘肽S转移酶p1(Gstp1),e磺基转移酶1b1(Sult1b1),f UDP葡糖醛酸糖基转移酶1a6 a / b (Ugt1a6a / b)。基因表达通过qPCR分析,并在十二指肠采用Rpl13a进行标准化。 在结肠中参考Hprt1和Rpl13a进行平均。 * P< 0.05; ** P< 0.01; 设置+Se为1,采用t-test分析方法进行分析。
此外,Burk及其同事提供了饮食中硒缺乏和Nrf2激活之间的第一个直接联系,比较了每公斤饲喂0.25毫克亚硒酸钠的完全缺硒或补充饮食的小鼠[9]。 在硒缺乏条件下,ARE驱动的报告基因活性以及GST和NQO1活性在野生型小鼠的肝脏中强烈增加,但在Nrf2 - / - 小鼠中没有变化。 目前的研究表明,边缘硒缺乏可导致II期酶的上调,强调了生物体对硒体内平衡变化的敏感程度。

图3 硒缺乏(-Se)小鼠与相对于不缺硒(+Se)小鼠的结肠(n = 9)和 十二指肠(n = 6)中硒蛋白的mRNA表达。 a 硫氧还蛋白还原酶1(Txnrd1), b谷胱甘肽过氧化物酶2(Gpx2),c硒蛋白W(Sepw1)。 基因表达通过qPCR分析,并在十二指肠采用Rpl13a进行标准化。 在结肠中参考Hprt1和Rpl13a进行平均。 * P< 0.05; ** P< 0.01; 设置+Se为1,采用t-test分析方法进行分析
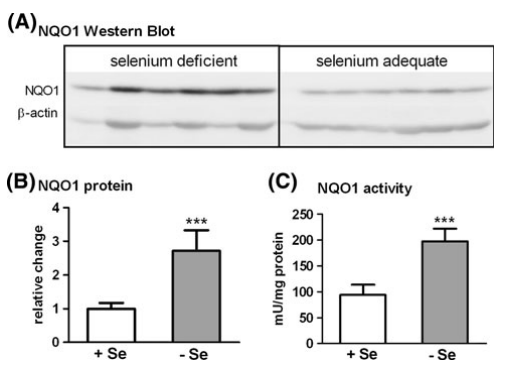
图4. 小鼠十二指肠中的NQO1蛋白表达和活性测试。 蛋白质测量通过Western Blot(a)方法,光密度分析与归一化采用b-actin(b)方法。 通过降低MTT方法(c)来测量绝对NQO1活性。 *** P< 0.001; 设置+Se为1,采用t-test分析方法进行分析,详情请参阅“材料和方法”
由于Nrf2靶基因表达不受肝脏中边缘硒缺乏的影响(表3),但在肠道中上调,可以得出结论,肠道对减少的硒供应更敏感。 在Nrf2 - / - 小鼠的肝脏中,大多数已知的Nrf2靶基因被下调,而主要用于解毒的酶在Keap1敲低小鼠中增加,其特征在于Nrf2激活增加[47]。 作者得出结论,Nrf2的肝脏激活对于解毒比对抗氧化防御更重要。 在目前的研究中,在十二指肠中观察到用于抗氧化防御的酶的明显上调。 由于减少的氧化还原状态维持肠上皮细胞增殖并防止早熟细胞凋亡[3],对抗氧化防御酶的更高需求可以解释目前的发现。
然而,也有相反的发现:当喂食0.01毫克硒/千克膳食或1毫克/千克时,解毒基因被硒缺乏下调[44]。 因此,硒含量相差100倍,而在本研究中,因子仅为2.相互矛盾的结果([13]中综述)可以解释为超营养饮食可以诱导II期酶更多 比硒缺乏有效。 以超营养饮食作为缺乏饮食的参考,净效应是硒缺乏的下调。
表3 肝脏中Nrf2靶基因和II期酶的qPCR结果

低硒含量和高硒含量影响的潜在机制似乎不同。 高浓度的某些硒化合物或代谢物可通过改变Keap1中的关键硫醇直接激活Nrf2途径。 反应性巯基也可以通过氧化来修饰[22,29],这种情况在硒缺乏中占主导地位[52,55]。 需要澄清中度缺硒小鼠中较高的氧化状态是否是诱导的Nrf2靶基因表达的原因。
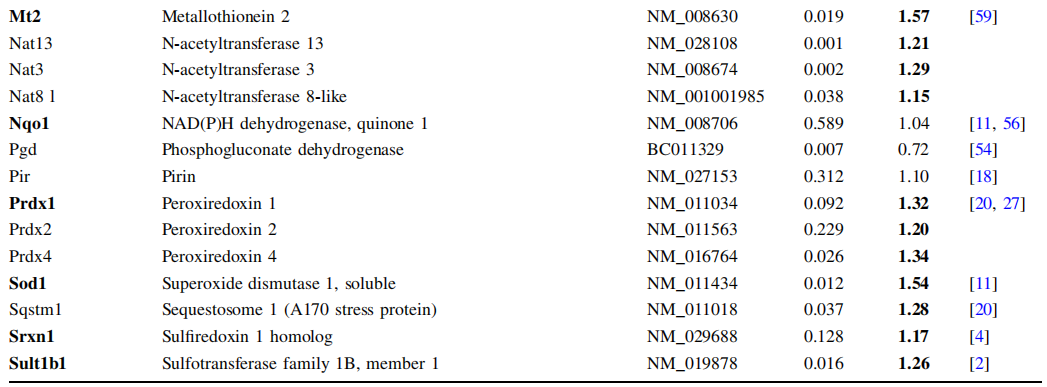
通过敲除肝细胞中Sec特异性tRNA(Trsp)的基因完全丧失所有硒蛋白,增强了几种II期酶的表达,表明至少一种硒蛋白通常抑制Nrf2通路的激活[50]。为了降低推定候选者的数量,Sengupta及其同事使用了第二只小鼠品系,其中Trsp突变的方式只有管家而非压力相关的硒蛋白表达。这些小鼠没有对II期酶的代偿性上调作出反应,这表明管家硒蛋白必须阻止Nrf2的活化[50]。由于硫氧还蛋白还原酶家族属于管家硒蛋白[10],因此分析了肝脏特异性TrxR1敲除对Nrf2途径的潜在激活。实际上,TrxR1敲除导致Nrf2的核积累和21个Nrf2靶基因的诱导[51]。 TrxR2,TrxR3或硫氧还蛋白的mRNA水平未受影响。因此,TrxR1似乎抵消Nrf2激活和/或用作Nrf2激活的关闭信号。然而,缺乏TrxR1的肝脏未显示氧化应激的证据,使潜在的机制不清楚[51]。
硒还影响编码非硒蛋白的多个基因的表达,这些基因可能参与边缘硒缺乏时Nrf2通路的特异性激活。 在之前的研究[28]中,我们发现Wnt通路被激活,表现为b-连环蛋白,Dvl2,Lef1和c-Myc的表达增强以及Gsk3β的下调。 Gsk3β是一种多功能丝氨酸/苏氨酸激酶,不仅参与糖原代谢和经典Wnt信号传导,而且还显示出使Nrf2通路失活[23,49]。 Fyn激酶被磷酸化,从而被Gsk3β激活。 反过来,Phospho-Fyn使Nrf2磷酸化,导致Nrf2的核输出[23]。 Gsk3β的活性降低,这可能是在先前研究中硒限制条件下从Gsk3β的下调推断出的,可能会降低活性Fyn的量,从而导致Nrf2的核积累。
边缘硒缺乏中Nrf2通路的激活与结直肠癌的发展有关。在炎症引发的结肠直肠癌发生模型中,与野生型小鼠相比,Nrf2 - / - 小鼠的发病率,多样性和肿瘤大小增加[25]。一方面,GST,SULT和UGT家族等II期酶参与异生物质的结合和排泄。加上抗氧化酶如过氧化物酶,硫化毒素,Sod1,Hmox1和金属硫蛋白等基因的上调,可以保护机体免受氧化损伤。另一方面,Nrf2激活也可能有助于增加药物输出,抗药性和肿瘤细胞存活[16,31]。因此,不能容易地确定Nrf2靶基因在硒缺乏中对结肠直肠癌的上调的结果。硒缺乏症中Wnt途径的激活指向低硒状态的癌症促进功能,并且与硒缺乏中较高的癌症发病率一致(在[46]中综述)。 Nrf2途径的激活是否是通过内源性防御系统的强化来补偿硒蛋白的损失的尝试,或者它是否已经有助于癌细胞的存活,这是进一步研究的挑战。
翻译:
缺硒条件下诱导Nrf2靶基因
摘要:在欧洲,硒摄入不足似乎是普遍存在的。因此,目前的研究主要集中在摄入不足的条件下基因表达的变化。之前,有研究采用微阵列分析方法,揭示了小鼠在充足硒营养或中度缺乏硒营养喂养条件下的结肠的几种途径的基因的变化。同时有研究发现,严重的硒缺乏会影响适应性反应的Nrf2调节的基因。由于先前的分析途径是采用不关联Nrf2靶基因的程序开展的,而通过qPCR手动选择和确认各自的基因表达变化,更具意义。qPCR显示在缺硒饮食条件下,诱导II相酶(Nqo1,Gsts,Sult1b1和Ugt1a6)和抗氧化酶(Hmox1,Mt2,Prdx1,Srxn1,Sod1和Gclc),这被认为是对硒蛋白损失的补偿。在十二指肠中观察到最强的效果,其中抗氧化酶的基因被优先上调。这些还包括硒蛋白TrxR1和GPx2的mRNA,它们能够在硒重新摄入时立即翻译表达。在先前的论文中,观察到的中度硒缺乏症中Gsk3β的下调,为Nrf2途径的激活提供了可能的解释,因为Gsk3β的抑制导致Nrf2的积累。关键词:硒;Nrf2; II相酶; 肠;氧化应激
引言
在哺乳动物中,必需的微量元素硒作为硒代半胱氨酸并入硒蛋白活性中心发挥作用。人类有25个基因编码硒蛋白[30],其中许多涉及氧化还原过程[35],包括谷胱甘肽过氧化物酶(GPx)[8]和硫氧还蛋白还原酶(TrxR)[14]。欧洲人口中存在的少量硒缺乏症可能伴随着轻微的氧化状态[45]。 硒蛋白的表达取决于细胞中硒的状态。但是,一些微阵列研究表明,硒的可用性也会影响非硒蛋白的表达(在[13]中进行了综述)。仍然缺乏基本机制。硒状态码经常影响抗氧化剂防御酶和II期系统的酶[13,44]。一方面,大剂量的硒刺激诸如谷胱甘肽S转移酶(GST)和NADPH:醌氧化还原酶(NQO1)等II期酶的活性。这种作用可以分别通过硒代半胱氨酸硒结合物[53]和二甲基二硒化物[58]来介导。另一方面,有强有力的证据证明在硒缺乏时会诱导抗氧化剂和II期酶的产生。早在1970年代末期,据报道血红素加氧酶1(HMOX1)[12]以及GST活性在缺硒的大鼠肝脏中有所升高[32]。硒缺乏症中抗氧化剂和II期酶的补偿性上调的想法进一步得到了小鼠的发现,该小鼠具有针对器官的靶向去除编码硒代半胱氨酸tRNA(Trsp)的基因。肝脏中硒蛋白的完全丧失导致了GSTPi,NQO1和HMOX1的诱导[52]。 参与细胞抗氧化剂防御和II期排毒的大多数蛋白质基因是由氧化还原和亲电敏感的转录因子NF-E2相关因子(Nrf2)调控的(在[16]中进行了综述),该因子与顺式作用的抗氧化剂或亲电反应元件(ARE / EpRE)[48,56]。 Nrf2-/-小鼠对化学诱导的毒性和肿瘤发生敏感[43]。另外,Nrf2-/-小鼠对氧化应激高度敏感[1],并具有较低的基础谷胱甘肽(GSH)水平[47]。在不受刺激的条件下,Nrf2保留在与Keap1结合的胞质溶胶中,Keap1充当基于Cul3的E3泛素连接酶的底物衔接子,并靶向Nrf2进行降解。靶基因的反式激活是响应活性氧(ROS)或亲电试剂而引起的,这些试剂修饰Keap1的易感硫醇基团,导致核Nrf2蛋白水平迅速提高(在文献[38]中进行了综述)。 Trsp和Nrf2的同时破坏消除了II期酶的诱导,从而验证了Nrf2在响应硒蛋白丢失中的作用[52]。与饲喂对照饮食的小鼠相比,饲喂缺硒饮食的ARE报告小鼠的ARE驱动的报告基因活性显著增强[9]。另外,在缺硒的野生型小鼠中观察到的GST和NQO1活性增加在Nrf2-/-小鼠中无法检测到[9]。这提供了饮食中硒缺乏与Nrf2激活之间的第一个直接联系。 基于这些结果,当前的研究集中在边缘缺硒饮食对Nrf2调控基因表达的影响上。贫硒饮食中小鼠的建议每日允许摄入量(RDA)为一半[28],反映出生理状况可能会因改变营养习惯或季节性食物变化而同样发??生。分析的目标器官是结肠和十二指肠的肠段,这是抵抗异种生物胁迫的第一道防线,还有肝脏,肝脏高度表达II期酶。先前研究[28]中获得的微阵列数据的途径分析未表明Nrf2途径受到影响,因为所用程序GenMAPP不包含该途径的MAPP(微阵列途径概况)。因此,手动鉴定了48个Nrf2靶基因和12个经典II期酶,并通过qPCR证实了有趣的候选基因。
材料和方法
动物实验
从先前报道的动物中获取组织样品[28]。对雄性C57BL / 6 J小鼠(3-4周龄)饲喂贫硒(0.086 mg Se / kg)或通过混合硒代蛋氨酸(Acros,比利时的盖尔(Geel)陷入贫困的人(德国的拉格(Altromin))。喂养6周后,麻醉的动物因颈脱位而被处死。冷冻固定在液氮中的血浆和组织储存在-80°C下。每组12只动物的硒状态以血浆硒含量和肝GPx活性为特征,发现在中等硒缺乏时分别降低至硒充足的13%和35%[28]。
RNA分离
在液氮冷却下将组织研磨。将20–30 mg的粉末悬浮在800μl的冷Trizol(Invitrogen,卡尔斯鲁厄,德国)中,并用组织裂解器(Qiagen,Hilden,德国)以30 Hz匀化2×2分钟。根据制造商的说明,使用Trizol方案分离RNA。用10 U RQ1 DNase(Promega,曼海姆,德国)消化基因组DNA,并通过苯酚-氯仿提取纯化RNA。使用NanoDrop ND-1000(Peqlab Biotechnologie GmbH,德国埃尔兰根)测量RNA浓度。
芯片分析
如前所述[28],使用Mouse 44 K芯片(Agilent Technologies,Böblingen,德国)进行芯片分析。Student’s t-test检验可鉴定出显著调控的基因(P <0.05)。 实时定量PCR 用150 fmol oligo(dT)15引物和180 U莫洛尼鼠白血病病毒逆转录酶(Promega)反转录RNA(3μg),总体积为45μl。
实时PCR
(Mx3005PTM QPCR系统,Stratagene,阿姆斯特丹,荷兰)使用SYBR Green I(Molecular Probes,Eugene,USA)作为荧光报告物,在25μl反应混合物中以1μl十倍稀释的cDNA一式三份进行。所有PCR反应的退火温度分别为60和62°C(表1)。 PCR产物的定量范围为1×103至1×109个拷贝的标准曲线。通过将至少一种引物置于PerlPrimer v1.1.14的外显子/内含子边界上,设计引物(表1,Sigma–Aldrich,Taufkirchen,德国)对cDNA具有特异性。参考基因Rpl13a和Hprt1的平均值用于在结肠和肝脏中qPCR结果的标准化,而Rpl13a仅用于十二指肠。
表1. 所用底物序列表
表2. 微阵列分析测量硒对结肠Nrf2靶基因作用
组织裂解液的制备
将10 mg磨碎的组织悬浮于含有2μl蛋白酶抑制剂混合物III(Calbiochem,Bad Soden,Germany)的250μl均质缓冲液(100 mM Tris-HCl,300 mM KCl,0.01%Triton X-100)中并用组织裂解器(Qiagen)在30 Hz下持续2×2分钟。裂解液在21,000×g和4°C下离心15分钟,然后根据Bradford [7]评估蛋白质含量。
蛋白质印迹
将等分试样(20μg蛋白/泳道)进行SDS-PAGE,并按[5]所述进行印迹。用兔抗人NQO1抗血清(1:3,000; ab34173,Abcam,剑桥,英国)检测到NQO1。通过兔抗人β-肌动蛋白抗血清(1:5,000; ab8229,Abcam)检测到的β-肌动蛋白用作内部对照。使用过氧化物酶偶联的抗兔抗体(1:50,000; Chemicon,霍夫海姆,德国)作为二抗。检测是在以SuperSignal West Dura(德国波恩的Perbio)为底物的Fuji LAS3000-CCD系统中完成的。用Aida / 2D光密度测定法4.0软件(Raytest,Straubenhardt,德国)对条带进行定量。
NQO1活动
根据[42]测量双香豆酚敏感的NQO1活性。在590 nm的微量滴定板吸光度读取器上追踪由甲萘醌介导的3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(MTT)的还原(Synergy 2,Biotek Instruments GmbH,Bad Friedrichshall,德国),将不同的填充量校正为1厘米的路径长度。将3μl裂解物与190μl反应缓冲液(25 mM Tris-HCl,pH 7.4,0.665 mg / l BSA,0.01%Tween 20,5μMFAD,1 mM葡萄糖-6-磷酸盐,30μM一起用于测定NADP,0.72 mM MTT,0.3 U / ml 6磷酸葡萄糖脱氢酶,50μM甲萘醌),总体积为250μl。 NQO1的活性计算为有和没有双香豆酚时MTT还原速率之间的差异,使用的还原MTT的消光系数在590 nm处估计为11,961(mol / l)-1×cm-1。
统计分析
比较两组,通过未配对的Student’s t-test(GraphPadPrism®5.0版,美国加利福尼亚州圣地亚哥)计算出显著差异。 P值<0.05被认为具有统计学意义。
结果
两个Nrf2硒蛋白靶基因GPx2和TrxR1是特殊情况,因为它们的翻译依赖于硒的可用性及其在层次结构中的排名。 高硒蛋白GPx2 [57]和TrxR1 [10]的mRNA在缺硒结肠中显著降低,GPx2受影响小于TrxR1。 相反,两种酶的mRNA在硒缺乏十二指肠中增强(图3a,b)。 硒缺乏时GPx2 mRNA的上调已经在细胞培养中显示[57]。 在限制硒条件下mRNA的稳定性使得在补充硒后能够在缺硒细胞中快速重新合成GPx2 [57]。 两种观察都可以作为硒蛋白质层次中GPx2高级别的证据。NQO1 RNA在缺硒十二指肠中的上调与NQO1蛋白表达和活性相关
在蛋白质和活性水平上进一步分析NQO1作为Nrf2靶基因的原型。 在十二指肠中,NQO1蛋白和活性在缺硒组中显著增强(图4),与增强的mRNA相关(图2a)。 在结肠中,蛋白质水平和活性都没有改变,表明NQO1 mRNA的轻微增加不会通过增加的翻译和活性反映出来。
讨论
本研究是动物实验的一部分,其中给小鼠喂食符合RDA的小鼠或硒含量降低至约50%的饮食[28]。虽然硒摄入量仅适度减少,但四种硒蛋白SelW,GPx1,SelH和SelM的RNA表达明显下调,Wnt途径的基因在结肠中上调[28]。对微阵列数据的人工分析表明,Nrf2靶基因和II期酶也受硒状态的调节;在结肠中表达的60个基因中,41个被上调占68%(表2)。通过qPCR证实了12种用于抗氧化防御和II期酶的所选基因的表达(图1和2)。 Nrf2靶基因在边缘硒缺乏中的一致上调是高度相关的,因为次优的硒供应也可能是由人类经常消耗的低硒饮食造成的[45]。边缘硒缺乏的建立与大多数其他研究高硒补充剂或严重缺硒的研究相反。
图1.硒缺乏(-Se)小鼠与相对于不缺硒(+Se)小鼠的结肠(n = 9)和 十二指肠(n = 6)中抗氧化酶的mRNA表达分析。 a血红素加氧酶1(Hmox1),b金属硫蛋白2(Mt2),c peroxiredoxin 1 (Prdx1), d sulfifiredoxin(Srxn1),e超氧化物歧化酶1(Sod1),f c-谷氨酰半胱氨酸合成酶,催化亚基(Gclc)。基因表达通过qPCR分析,并在十二指肠采用Rpl13a进行标准化。 在结肠中参考Hprt1和Rpl13a进行平均。 * P< 0.05; ** P< 0.01; 设置+Se为1,采用t-test分析方法进行分析
图2.硒缺乏(-Se)小鼠与相对于不缺硒(+Se)小鼠的结肠(n = 9)和 十二指肠(n = 6)中II相酶的mRNA表达。NADPH:醌氧化还原酶(Nqo1), b谷胱甘肽S转移酶a1 / a2(Gsta1 / a2),c谷胱甘肽S转移酶m1(Gstm1),d谷胱甘肽S转移酶p1(Gstp1),e磺基转移酶1b1(Sult1b1),f UDP葡糖醛酸糖基转移酶1a6 a / b (Ugt1a6a / b)。基因表达通过qPCR分析,并在十二指肠采用Rpl13a进行标准化。 在结肠中参考Hprt1和Rpl13a进行平均。 * P< 0.05; ** P< 0.01; 设置+Se为1,采用t-test分析方法进行分析。
此外,Burk及其同事提供了饮食中硒缺乏和Nrf2激活之间的第一个直接联系,比较了每公斤饲喂0.25毫克亚硒酸钠的完全缺硒或补充饮食的小鼠[9]。 在硒缺乏条件下,ARE驱动的报告基因活性以及GST和NQO1活性在野生型小鼠的肝脏中强烈增加,但在Nrf2 - / - 小鼠中没有变化。 目前的研究表明,边缘硒缺乏可导致II期酶的上调,强调了生物体对硒体内平衡变化的敏感程度。
图3 硒缺乏(-Se)小鼠与相对于不缺硒(+Se)小鼠的结肠(n = 9)和 十二指肠(n = 6)中硒蛋白的mRNA表达。 a 硫氧还蛋白还原酶1(Txnrd1), b谷胱甘肽过氧化物酶2(Gpx2),c硒蛋白W(Sepw1)。 基因表达通过qPCR分析,并在十二指肠采用Rpl13a进行标准化。 在结肠中参考Hprt1和Rpl13a进行平均。 * P< 0.05; ** P< 0.01; 设置+Se为1,采用t-test分析方法进行分析
图4. 小鼠十二指肠中的NQO1蛋白表达和活性测试。 蛋白质测量通过Western Blot(a)方法,光密度分析与归一化采用b-actin(b)方法。 通过降低MTT方法(c)来测量绝对NQO1活性。 *** P< 0.001; 设置+Se为1,采用t-test分析方法进行分析,详情请参阅“材料和方法”
由于Nrf2靶基因表达不受肝脏中边缘硒缺乏的影响(表3),但在肠道中上调,可以得出结论,肠道对减少的硒供应更敏感。 在Nrf2 - / - 小鼠的肝脏中,大多数已知的Nrf2靶基因被下调,而主要用于解毒的酶在Keap1敲低小鼠中增加,其特征在于Nrf2激活增加[47]。 作者得出结论,Nrf2的肝脏激活对于解毒比对抗氧化防御更重要。 在目前的研究中,在十二指肠中观察到用于抗氧化防御的酶的明显上调。 由于减少的氧化还原状态维持肠上皮细胞增殖并防止早熟细胞凋亡[3],对抗氧化防御酶的更高需求可以解释目前的发现。
然而,也有相反的发现:当喂食0.01毫克硒/千克膳食或1毫克/千克时,解毒基因被硒缺乏下调[44]。 因此,硒含量相差100倍,而在本研究中,因子仅为2.相互矛盾的结果([13]中综述)可以解释为超营养饮食可以诱导II期酶更多 比硒缺乏有效。 以超营养饮食作为缺乏饮食的参考,净效应是硒缺乏的下调。
表3 肝脏中Nrf2靶基因和II期酶的qPCR结果
低硒含量和高硒含量影响的潜在机制似乎不同。 高浓度的某些硒化合物或代谢物可通过改变Keap1中的关键硫醇直接激活Nrf2途径。 反应性巯基也可以通过氧化来修饰[22,29],这种情况在硒缺乏中占主导地位[52,55]。 需要澄清中度缺硒小鼠中较高的氧化状态是否是诱导的Nrf2靶基因表达的原因。
通过敲除肝细胞中Sec特异性tRNA(Trsp)的基因完全丧失所有硒蛋白,增强了几种II期酶的表达,表明至少一种硒蛋白通常抑制Nrf2通路的激活[50]。为了降低推定候选者的数量,Sengupta及其同事使用了第二只小鼠品系,其中Trsp突变的方式只有管家而非压力相关的硒蛋白表达。这些小鼠没有对II期酶的代偿性上调作出反应,这表明管家硒蛋白必须阻止Nrf2的活化[50]。由于硫氧还蛋白还原酶家族属于管家硒蛋白[10],因此分析了肝脏特异性TrxR1敲除对Nrf2途径的潜在激活。实际上,TrxR1敲除导致Nrf2的核积累和21个Nrf2靶基因的诱导[51]。 TrxR2,TrxR3或硫氧还蛋白的mRNA水平未受影响。因此,TrxR1似乎抵消Nrf2激活和/或用作Nrf2激活的关闭信号。然而,缺乏TrxR1的肝脏未显示氧化应激的证据,使潜在的机制不清楚[51]。
硒还影响编码非硒蛋白的多个基因的表达,这些基因可能参与边缘硒缺乏时Nrf2通路的特异性激活。 在之前的研究[28]中,我们发现Wnt通路被激活,表现为b-连环蛋白,Dvl2,Lef1和c-Myc的表达增强以及Gsk3β的下调。 Gsk3β是一种多功能丝氨酸/苏氨酸激酶,不仅参与糖原代谢和经典Wnt信号传导,而且还显示出使Nrf2通路失活[23,49]。 Fyn激酶被磷酸化,从而被Gsk3β激活。 反过来,Phospho-Fyn使Nrf2磷酸化,导致Nrf2的核输出[23]。 Gsk3β的活性降低,这可能是在先前研究中硒限制条件下从Gsk3β的下调推断出的,可能会降低活性Fyn的量,从而导致Nrf2的核积累。
边缘硒缺乏中Nrf2通路的激活与结直肠癌的发展有关。在炎症引发的结肠直肠癌发生模型中,与野生型小鼠相比,Nrf2 - / - 小鼠的发病率,多样性和肿瘤大小增加[25]。一方面,GST,SULT和UGT家族等II期酶参与异生物质的结合和排泄。加上抗氧化酶如过氧化物酶,硫化毒素,Sod1,Hmox1和金属硫蛋白等基因的上调,可以保护机体免受氧化损伤。另一方面,Nrf2激活也可能有助于增加药物输出,抗药性和肿瘤细胞存活[16,31]。因此,不能容易地确定Nrf2靶基因在硒缺乏中对结肠直肠癌的上调的结果。硒缺乏症中Wnt途径的激活指向低硒状态的癌症促进功能,并且与硒缺乏中较高的癌症发病率一致(在[46]中综述)。 Nrf2途径的激活是否是通过内源性防御系统的强化来补偿硒蛋白的损失的尝试,或者它是否已经有助于癌细胞的存活,这是进一步研究的挑战。
本文由福山生物整理翻译,转载请注明出处。