






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
萝卜硫素通过Nrf2信号传导预防年龄相关的心脏和肌肉功能障碍
发表于:2020-10-29 作者:admin 来源:本站 点击量:11235
摘要:
年龄相关的线粒体功能障碍和氧化损伤是多种健康问题的主要原因,包括肌肉减少症和心血管疾病(CVD)。尽管Nrf2(一种调节细胞保护性基因表达的转录因子)在肌肉病变中的作用仍不清楚,但它在肌肉减少症和CVD中均显示出有益的特性。萝卜硫素(SFN)是一种激活Nrf2相关细胞保护基因的天然化合物,可对包括CVD在内的多种疾病提供保护,并且正在进行从预防癌症到降低胰岛素抵抗的各个阶段临床试验。这项研究旨在确定SFN是否可以预防与年龄相关的心脏和骨骼肌功能丧失。对2个月大和21至22个月大的小鼠进行常规啮齿动物饮食或补充SFN的饮食12周。在研究完成时,检测了骨骼肌和心脏功能、线粒体功能和Nrf2活性。我们的研究表明,与年轻组小鼠相比,老年对照组小鼠的Nrf2活性和线粒体功能显著下降,并且骨骼肌和心脏功能丧失。在老年小鼠中,SFN恢复了Nrf2活性、线粒体功能、心脏功能、运动能力、葡萄糖耐量以及骨骼肌卫星细胞的激活/分化。我们的结果表明,与年龄相关的Nrf2信号传导活性下降和与之相关的线粒体功能障碍可能与年龄相关疾病的发展有关。因此,通过SFN恢复Nrf2活性和内源性细胞保护机制可能是一种安全有效的策略,可以防止由于衰老而引起的肌肉和心脏功能障碍。
关键词:心脏功能、线粒体功能障碍、Nrf2、氧化应激、肌肉减少症、萝卜硫素
1. 前言
衰老是受内在和外在因素共同调节的复杂生物过程。衰老的机制包括活性氧(ROS)、端粒缩短和激素变化的作用。 年龄相关的线粒体功能障碍和氧化损伤是多种健康问题的主要原因,包括肌肉减少症和心血管疾病(CVD)(Miller et al.,2012)。已经评估了几种化学和功能不同的ROS和抗氧化剂产品的清除剂缓解年龄引起的肌肉丢失的能力(Barrera等人,2018),但收效甚微。它们缺乏益处的主要原因可能包括细胞系统中生物利用度低和/或对氧化剂和亲电试剂的清除能力低。因此,促进内在抗氧化剂和细胞保护途径的活性可能代表了一种对抗老年人肌肉功能丧失的更有效策略。
对于细胞适应氧化应激和亲电子应激至关重要的酶(和其他蛋白质)的转录调控是由抗氧化剂反应性DNA元件(ARE)控制的,也称为亲电反应元件(Suh等人,2004)。细胞对亲电应激反应的主要调节因子是转录因子Nrf2(核红细胞-2-p45相关因子-2),被证明对解毒基因活性至关重要(Itoh等,2004),包括在哺乳动物的心脏细胞中以及心血管系统的其他组成部分(Dastani等,2012)。尽管Nrf2在肌病中的作用尚不明确,但其下调与肌肉减少症和CVD相关(Gounder等,2012)。在生理条件下,Nrf2通过其阻遏因子,kelch样ech结合蛋白1 (Keap1)结合在细胞质中。Keap1调节亲电体对Nrf2的降解(Itoh等,2004)。亲电物,包括天然化合物萝卜硫素(SFN),通过Keap1的硫醇修饰激活抗氧化和细胞保护途径,使其在细胞质中与Nrf2分离。释放后,Nrf2积聚在细胞核中,与小的Maf蛋白异源二聚体,并通过其AREs激活靶基因的转录(Cao等人,2006)。 Nrf2靶基因编码参与细胞保护反应的蛋白质,包括细胞抗氧化剂和抗亲电子活性以及重要的细胞抗氧化剂谷胱甘肽(GSH)的生物合成。迄今为止,亲电代谢中编码酶的所有已知基因以及众多抗氧化酶均在基础表达水平受到调控,并经常被Nrf2诱导(Dinkova-Kostova&Abramov,2015)。
产生过量ROS的线粒体缺陷对细胞有害,并经常通过自噬/线粒体途径清除,然后诱导线粒体生物发生,以试图恢复线粒体的质量和功能(DinkovaKostova&Abramov,2015)。Nrf2通过直接激活保护机制来维持氧化应激期间的线粒体形态和完整性(Dinkova-Kostova&Abramov,2015)。
SFN是十字花科蔬菜中的一种植物化学物质,是一种无毒化合物,因其在不同疾病模型中的抗衰老、抗癌、抗糖尿病、抗微生物和化学预防活性而被公认(Kensler等,2013)。增强的Nrf2信号传导和随后的细胞保护性基因激活被认为是SFN作用的主要机制(Bai等人,2013)。SFN已显示出减少小胶质细胞介导的神经炎症的潜力,并可以减轻神经性行为缺陷并减轻阿尔茨海默氏病模型小鼠的αβ负担(Uddin等,2020)。SFN改善肌肉功能,改善病理学,保护营养不良的肌肉并减轻肌肉发炎(Sun等,2015)。SFN还通过Akt/Foxo信号传导抑制地塞米松诱导的肌管肌肉萎缩(Son等人,2017)。此外,SFN可以通过促进抗氧化剂机制修复与年龄有关的心血管疾病中的血管平滑肌细胞功能障碍,并防止皮肤衰老(Sedlak等人,2018)。目前,一些正在进行的临床前和临床试验研究了其对癌症、胰岛素抵抗、精神分裂症和自闭症的作用(Kensler等,2013; K. Singh等,2014)。
因为已知SFN可以激活Nrf2并减轻氧化损伤,所以在这项研究中,我们研究了SFN处理是否可以恢复老年小鼠的Nrf2活性、线粒体功能以及骨骼肌和心脏功能。我们还评估了SFN处理和对照小鼠的骨骼肌和心脏中的转录组改变。在这项研究中,我们为上调Nrf2活性作为一种缓解年龄诱导的肌肉和心脏功能障碍的新治疗策略奠定了基础。
2. 结论
2.1 SFN提高生存率,但不影响动物体重
在21-22个月大时,将雄性C57BL / 6小鼠置于含有SFN的饮食或对照饮食(每组n = 20小鼠)中,共12周。在SFN给药期间,对照组中有6只小鼠死亡。另一方面,SFN饮食组中的所有小鼠均存活到实验结束,直到它们达到24至25个月大时为止(图1a)。膳食补充SFN并不会影响年轻或成年小鼠的体重。正如预期的那样,在两个饮食组中,年轻小鼠的体重增长都快于老年小鼠(图1b)。另一方面,补充SFN会影响进水量(图1c)。与对照组饮食相比,SFN饮食喂养的年轻小鼠的饮水量显著降低。在年长的动物群中也观察到了同样的情况。但是,食物摄入量不受影响(图1d)。此外,补充SFN不会影响年轻和老年小鼠的血液学参数(表S1)。
2.2 SFN可以降低老年小鼠的空腹血糖并改善其葡萄糖耐量
在SFN饮食中,年轻小鼠的空腹血糖水平不高于年轻对照小鼠(图1e)。另一方面,饲喂SFN日粮的老年小鼠与老年对照组小鼠(102±9 mg / dl)相比,葡萄糖水平显著降低(平均±SD:77±16 mg / dl)。在两个饮食组中,葡萄糖水平都在C57BL / 6小鼠的正常范围内或以下(Surwit等,1988)。由于对照组和治疗组动物的空腹血糖水平差异不大,因此进行了腹膜内葡萄糖耐量试验。SFN处理不影响年轻动物对葡萄糖给药的反应(图1f)。但是,与对照组相比,在接受饮食控制的老年小鼠中,注射葡萄糖后血糖水平显著增加。此外,SFN治疗逆转了这种血糖升高的情况,并改善了老年动物的葡萄糖耐量(图1f)。
2.3 SFN可改善老年小鼠的运动能力
我们测试了SFN治疗是否会改善肌肉力量和运动能力。为此,我们研究了接受对照或SFN饮食喂养的年轻和老年小鼠,进行了不随意的体育锻炼来测试其肌肉力量。首先,我们通过将它们固定在粗线上来测试它们的抓地力。用对照饮食喂养的年轻和老年小鼠能够分别平均维持54±5和39±5 s。用SFN饮食的年轻和老年小鼠能够保持更长的时间,分别为64±13和86±12 s(图2a)。令人惊讶的是,喂SFN的老年小鼠比喂SFN的年轻小鼠能坚持更长的时间。此外,我们比较了对照组和SFN处理的年轻小鼠和老年小鼠的运动能力,方法是让它们在电动,速度可控的模块化跑步机系统上运行(Handschin等,2007)。与年轻的同龄小鼠相比,接受对照饮食的老年小鼠的运动能力较低(图2b)。饲喂富含SFN的饮食可以显著改善老年小鼠的运动能力。由SFN喂食的老年老鼠在跑步机上的表??现与年轻动物相似。
2.4 SFN增加了骨骼肌干细胞的数量及其功能
从下肢的EDL肌肉分离的肌纤维培养72小时。固定了卫星细胞衍生的成肌细胞,并对Pax7和MyoD进行了免疫染色,以此作为卫星细胞增殖和分化的指标(图S1-S8)。大部分卫星细胞表达Pax7,甚至更多表达MyoD(图2c)。Pax7在SKM干细胞中的表达不影响MyoD表达,并有助于SKM的生长和再生(Zammit等,2006)。 在SFN饮食的年轻和老年小鼠中,Pax7-(图2c)和MyoD(图2d)阳性卫星细胞子代的比例显著高于其年龄匹配的对照组。SFN治疗可能会增加新的卫星细胞形成,从而满足肌肉稳态的常规需求,或者可能是偶发性的肥大或修复需求。
2.5 SFN减少了SKM老化,氧化和凋亡的标志
组织学显示,使用SFN饮食2个月后,载玻片的横截面积更多地被肌纤维占据(图S9)。同样,肌肉肌抑制素(SKM质量的负调节剂)的染色也较少(图S10)。最后,我们发现有证据表明SFN可以降低8OHdG(一种氧化的标志物)(图S11)和Tunel染色(一种凋亡的替代标志物)(图S12)。
2.6 SFN治疗老年小鼠也可改善心脏功能
已知心脏/呼吸功能下降会限制老年人的运动能力(Farkhooy et al。,2018)。心脏衰老是一个内在的过程,具有深刻的细胞和分子变化,可导致心脏功能受损(Vigorito&Giallauria,2014)。我们检查了SFN是否能恢复老年小鼠的心脏功能。与对照饮食的年轻小鼠相比,对照饮食的老小鼠射血分数(61.0±1.0%)、短轴缩短率(32.1±0.7%)和每搏输出量(32.8±9.2 µl)均降低(图3a-c)。另一方面,补充SFN可以改善老年小鼠的射血分数(76.0±1.4),短轴缩短率(44.2±1.3%)和每搏输出量(51.6±11.3 µl)。尽管SFN还可以显著增加年轻小鼠的这些参数,但对老年小鼠的射血分数和分数缩短的影响更大。补充SFN对年轻小鼠的心输出量无明显影响,而老年小鼠的心输出量则有明显改善(图3d)。结果,SFN喂养的老年小鼠的心输出量与年轻对照组的相似。我们得出的结论是,SFN喂养的老年小鼠对与年龄相关的心脏功能丧失具有抵抗力。
2.7 SFN保护衰老心脏中的线粒体超微结构
线粒体是高能量需求型心肌细胞的主要能源,占据其细胞体积的40%以上(约5,000个线粒体/心肌细胞)。心肌细胞功能严重依赖于这些细胞器的健康状况(Strom等,2016)。此外,与年龄相关的线粒体功能障碍可能是由于氧化损伤导致线粒体超微结构的变化所致(Vays等,2014)。为了确定SFN是否能预防与年龄相关的心脏线粒体超微结构变化,我们使用透射电子显微镜比较了对照组和SFN喂养的老年小鼠心脏之间的线粒体形态。结果表明,SFN保护线粒体免于与年龄相关的嵴紊乱、部分嵴溶解和基质电子密度降低的影响(图4a)。由于参与氧化磷酸化的酶位于线粒体内膜上,嵴的表面积和数量通常与细胞的代谢活性相关(Arismendi-Morillo,2011)。为了进一步验证在用SFN处理的老年小鼠心脏中观察到的电子转运链(ETC)复合物活性的增加/保护,我们用针对复合物I(NDUFS3-a核DNA编码亚基)和复合物III的蛋白质的抗体进行了免疫染色(CORE2-线粒体编码的亚基)。与年龄匹配的未治疗动物的心脏相比,SFN处理后衰老的心脏中的线粒体复合物I和III亚基蛋白表达增加(图4b)。这些数据表明,SFN可能维持线粒体功能的一种机制是诱导线粒体蛋白表达。另外,在老年小鼠的心肌线粒体中发现的蛋白质表达下降可能是由于氧化损伤。左心室组织的免疫组织化学分析显示,SFN喂养的老年小鼠具有较低水平的硝基酪氨酸蛋白加合物(氧化应激的标志物)(图4c)。这些作用可能有助于增加老年小鼠心脏的自噬,并通过SFN减轻自噬,如LC3-II在老年小鼠中的出现及其在SFN治疗后的消失所证明的(图S13)。
2.8 SFN改善老年小鼠的线粒体功能
心脏组织的线粒体特别丰富,可以满足其高代谢需求。线粒体呼吸复合物的破坏可导致氧化剂的产生,ATP耗竭和心肌细胞功能障碍。线粒体功能一直被认为在衰老过程中会下降,从而导致氧化应激增加(Hebert等,2015)。我们通过高分辨率呼??吸测定法测量了心脏和SKM组织的线粒体功能。与年轻的小鼠心脏相比,老年人的ETC活性低约30%,SFN保护ETC免受这种与年龄相关的氧通量下降(图5a)。此外,在SFN喂养的动物中ETC复合物I,I + II和最大呼吸的活性增加,但这种增加仅在老年小鼠中才显著。SFN喂养动物的SKM的线粒体中也发现了这种类似趋势,但并未达到显著水平(图5b)。
2.9 SFN恢复老年小鼠心脏和SKM中的Nrf2活性
除了对线粒体的影响外,Nrf2功能与年龄相关的下降已有充分的文献记载(Gounder等,2012),但尚不清楚SFN是否能在老年人群中恢复Nrf2的活性。最近,我们报道Nrf2活性随SFN处理而增加,并且是保护心脏免受诸如阿霉素和电离辐射之类的氧化剂伤害的关键(Boerma等,2015; P。Singh等,2015)。因此,我们比较了Nrf2 ARE结合活性在年轻和老年小鼠的心脏和SKM中的作用。与年龄匹配的对照组相比,SFN喂养的年轻和老年小鼠的心脏和SKM的核提取物中Nrf2活性均上调(图6)。
2.10心脏和骨骼肌的qRT-PCR结果进一步描述了SFN发挥作用的潜在机制
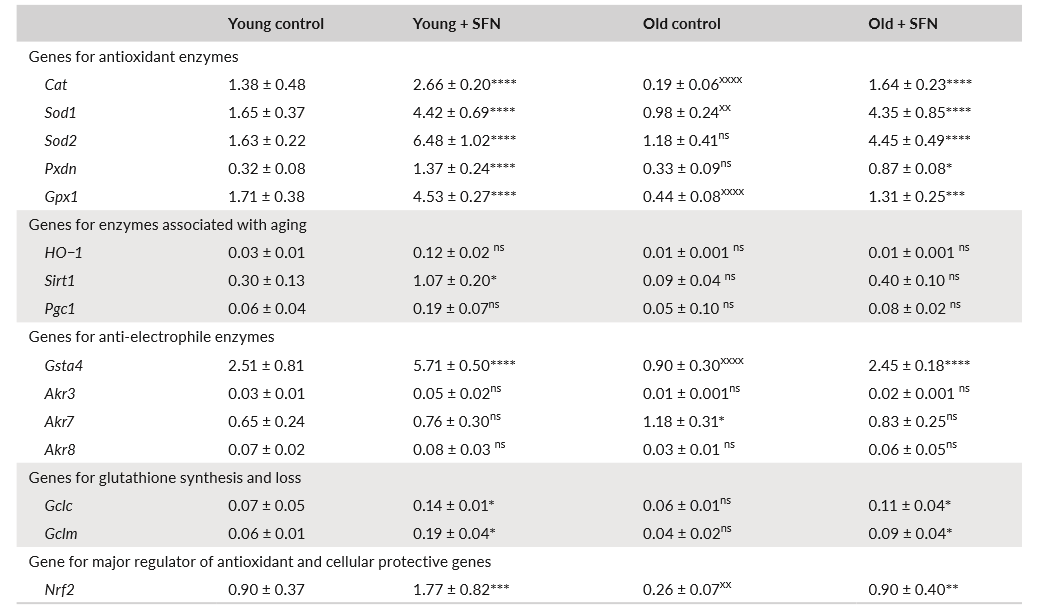
为了进一步阐明SFN介导的小鼠心脏和SKM功能衰老增强的机制,我们检查了许多与氧化剂和亲电子代谢相关的基因的表达。选定的基因参与抗氧化剂,抗亲电子活性,哺乳动物的寿命以及谷胱甘肽的合成和损失(表1和2),所有这些基因均受Nrf2的调控(Hayes&Dinkova-Kostova,2014)。我们检查了过氧化氢酶,Sod1,Sod2,HO1,Pxdn,Gpx1,HO-1,Gsta4,Akr3,Akr7,Akr 8,Sirt1,Pgc1,Gclc,Gclm和Nrf2的心脏和SKM转录水平(全表见表1基因)。在对Gclc,Gclm,Gsta4,Nqo-1,Ho-1和Sod2启动子区域进行的计算机或体外分析中,发现了Nrf2的共有ARE结合位点(Tonelli et al。,2018)。
与饲喂对照饮食的小鼠相比,SFN饮食喂养的年轻和老年小鼠心脏中的许多抗氧化剂和抗亲电子基因的心脏转录水平显著增加(表1)。新陈代谢的这种变化可以解释为什么含有SFN的饮食具有心脏保护作用,并能增强运动能力。在对照组中,大多数检查的基因以较低的水平表达。与SFN喂养的动物相比,老年对照动物产生的较低水平的抗氧化剂和抗电子防御机制可能有助于心脏的氧化损伤。在用SFN日粮喂养的动物中,某些转录本的水平,特别是Sod1,Sod2,Cat和Nrf2,在老小鼠的心脏中降低,并且在日粮中被SFN基本上恢复了。Akr3证明了老年小鼠心脏的增加,并且在饮食中通过SFN得以部分恢复。
表2给出了来自相同小鼠的SKM的类似实验的结果。在这种情况下,这些基因中的更多基因在老年小鼠中被下调,并且在饮食中被SFN很大程度上恢复:Cat,Sod1,Gpx1,Gsta4和Nrf2。尽管Sod2,Pxdn,Gclc和Gclm不受衰老的影响,但在老年小鼠的饮食中,它们也被SFN上调。总之,这些结果表明Nrf2途径是在SFN治疗后在老年小鼠中诱导的,并增强了SKM中的保护机制,可能甚至超过心脏。总而言之,这些结果表明Nrf2途径是在SFN处理后在年轻和老年小鼠中诱导的,并增强了保护机制。
3. 讨论
SFN可以延长老年小鼠的寿命,降低其患糖尿病的可能性,并提高其运动能力。运动能力增加的部分原因当然是由于SKM功能增强,但某些原因可能是由于保留了心脏功能。此外,在研究期间比较了使用SFN或对照饮食的成年小鼠的存活曲线,显示了显著的保护作用,从采用SFN饮食的老年动物的死亡率中得出结论(对数秩(Mantel-Cox)检验,p <.0087; Gehan–Breslow–Wilcoxon检验,p <.0089)。
我们使用年轻和老年的C57BL / B6J小鼠进行了SFN饮食研究,因为该小鼠寿命相对较长,并且死因各异,但肿瘤发生率低(Treuting等,2008)。先前的研究表明,SFN治疗会降低以高脂饮食或以高脂高蔗糖饮食喂养的肥胖小鼠为食的小鼠的体重增加和食物摄入,并改变其代谢参数(Shawky等人,2016; Shawky&Segar, 2018)。使用更正常的饮食,我们发现SFN对体重或食物摄入量没有影响,尽管我们确实发现用SFN喂养的老年小鼠的水摄入量减少了。我们推测增加的肌肉质量可能弥补了水摄入减少所引起的体重差异。小鼠对血液学参数无统计学显著影响(表S1)。使用SFN饮食的老年小鼠的空腹血糖水平比老年对照动物的空腹血糖水平稍低,但仍保持在正常范围内。如图1f所示,IPGTT在年轻小鼠中是正常的,但是与SFN饮食中的老年小鼠相比,对照饮食中的老年小鼠中胰岛素抵抗变得明显。个别年龄较大的小鼠(对照组和SFN饮食)对胰岛素的反应之间的差异表明,采用对照饮食的老年小鼠开始出现代谢综合征和病理。这些结果表明,SFN饮食可能对这些老年小鼠的胰岛素抵抗具有有益的作用。
目前的研究结果包括,在使用SFN饮食的老年小鼠中,跑步机的运动能力和对前肢握力的反应明显改善。饲喂SFN日粮的老年小鼠的跑步机运动能力大约是饲喂对照饮食的小鼠的1.5倍。当比较前肢的握力时,用SFN喂养的老年小鼠挂在电线上的时间是使用对照饮食的老年小鼠的两倍以上。用SFN喂养的老年小鼠的表现甚至比年轻的小鼠还要好。用SFN喂养的小鼠的EDL肌纤维中Pax7和MyoD阳性卫星细胞的子代增加,表明老年小鼠的肌肉再生得到改善。总体而言,我们已经证明SFN饮食干预可以改善老年小鼠的运动表现,抓地力和增加肌肉干细胞的形成,这表明SFN可能是减少衰老人群肌肉减少症的一种新型治疗方法。
SFN还对老年小鼠的心脏功能产生治疗作用。我们的结果表明,SFN治疗可显著改善老年小鼠的心脏射血分数,缩短分数,中风量和心输出量,使老年小鼠的静息心功能达到年轻小鼠的水平。这些特定的小鼠似乎比大多数其他研究的小鼠品系更容易发生收缩功能障碍,这些品系主要患有舒张功能障碍(de Lucia等人,2019)。然而,有报道说衰老小鼠的左心室射血分数降低导致收缩功能障碍(Han等,2020; Quarles等,2020),并且我们以前已经证明SFN可以防止阿霉素诱导的氧化应激左心室射血分数和分数缩短的异常(Benes et al。,2013; Bose et al。,2018; Singh et al。,2015)。
这项研究有几个重要的局限性。它仅在雄性小鼠中进行;因此,性别差异是未知的。此外,这项研究是在相对较小的样本量下进行的,仅使用了两个年龄组,并且持续时间相对较短(12周)。我们尚未完全描述SKM与心脏受SFN影响的机制的差异。最后,将结果转换为人类是未知的,尤其是在心力衰竭方面。然而,很明显,老年人的骨骼肌和心肌功能均下降,并且极大地降低了生活质量。在这种情况下,SFN可能是一种有希望的策略,可以减轻这些组织的衰老过程。
衰老心脏中ROS的产生增加可导致心肌细胞的坏死和凋亡以及成纤维细胞的增殖以及胶原蛋白的过量生成和纤维化的发展,以及线粒体的损??害;这些事件随后导致心脏重塑和功能障碍(Dai等,2014)。不幸的是,心脏中的抗氧化剂和亲电子保护性酶水平较低,使其特别容易受到自由基的损害(Singh等,2015)。年龄相关的线粒体功能障碍是衰老的重要部分。这些能量源能够自我复制,但随着年龄的增长,会通过各种机制(包括氧化/亲电子应力的持续恶性循环)而失效。为了进一步确定SFN在衰老过程中在线粒体功能和形态中的作用,我们评估了线粒体的超微结构,关键的电子转运链复合物蛋白表达以及小鼠心脏中氧化应激的标志物以及心脏和SKM中的线粒体呼吸复合物。氧化剂和亲电子试剂的增加,导致衰老期间的Nrf2活性和Nrf2响应基因,修饰了参与能量代谢的几种线粒体蛋白,并导致嵴的混乱。从我们对心脏和SKM活组织检查的高分辨率呼吸测定研究中获得的数据表明,衰老会损害复杂的I,I&II和最大呼吸功能,但这种影响在心脏中更为明显。由SFN喂养的年老小鼠的线粒体复杂活动的心脏已完全恢复,接近年轻对照组的水平。由于这些活动在老年动物的心脏活检中严重降低,因此恢复具有重要意义。喂食老年动物SFN后,CORE-2和NDUFS3表达增加,线粒体嵴混乱、部分嵴溶解、电子透明基质以及硝基酪氨酸蛋白加合物的水平降低,说明也可能有助于改善线粒体功能以及通过Nrf2依赖性细胞防御激活的SFN保护作用。我们认为通过SFN药理激活Nrf2可能会通过增强2相排毒和抗氧化酶活性并保留电子传输链(ETC)复杂功能来消除衰老过程中产生的过量ROS。由于在线粒体较少的SKM中,其对线粒体功能的影响不太明显,因此SFN可能对SKM发挥其他额外作用。
细胞中的累积氧化损伤是细胞衰老和衰老的基本特征。与年龄相关的氧化应激会促进心肌纤维化,左心室硬化的进行性增加以及心脏功能障碍,并且可能是导致肌肉无力和肌肉减少症的因素之一。与年龄相关的Nrf2功能障碍可降低抗氧化剂和抗亲电子酶的活性以及线粒体功能,导致体内稳态失调,并损害细胞的修复和再生能力。我们以前已经在体外和体内证明了SFN保护和增强Nrf2信号传导,通过激活细胞保护性基因以抵抗辐射和阿霉素引起的心脏毒性以及其他相关疾病(Chapple等,2012)。我们最近证明,SFN增强了小鼠心脏中的Nrf2信号传导和细胞保护性基因活性,从而保护免受氧化损伤(Singh等人,2015)。
在本研究中,我们研究了饮食中的SFN是否能激活两个最重要的功能组织SKM和心脏中的Nrf2活性和响应基因。我们确定了喂食SFN或对照饮食的年轻和老年小鼠的SKM和心脏中活性核Nrf2的量。SKM和SFN日粮喂养的年轻和老年小鼠的心脏显示ARE结合活性显著增加,这与对照饮食中老年小鼠中观察到的活性显著降低相反。老年对照组小鼠中Nrf2活性的下降清楚地表明,SKM和心脏(已经表达了低水平的保护酶)无法保护自己免受氧化和亲电攻击并正常运行。因此,在这项研究中,我们首次证明了衰老的心脏和SKM中氧化应激的增加与Nrf2调节异常引起的保护性反应失败有关,并且可以通过让老年小鼠(21到22个月大的小鼠)服用SFN来减轻这一过程。
检测了对照组和SFN饲料喂养小鼠心脏(A)和SKM(B)中负责表达抗氧化、抗亲电性和谷胱甘肽合成途径酶的基因转录水平。(A)在心脏中,过氧化氢酶、Sod1、Sod2和Nrf2与衰老小鼠的功能降低以及SFN的恢复表现出最佳的相关性。此外,Akr3在老年小鼠中增加,SFN减少了这种增加。Sod1和Sod2保护线粒体基质中的线粒体复合物和活性酶,并维持体内稳态和线粒体完整性(Piao等人,2010)。(B)在SKM中,可能的因素众多:过氧化氢酶、Sod1、Gpx1、Gsta4和Nrf2。心脏和SKM中受影响的基因之间只有部分重叠并不奇怪,特别是考虑到衰老和SFN对SKM中线粒体的影响较小。过氧化氢酶和Sod1可能在线粒体水平上发挥某些作用,但也可以增加抗氧化剂和抗电子防御机制的水平,并有助于降低脂质过氧化产物和4-HNE的浓度,从而恢复老年小鼠SKM的功能。酶促清除超氧化物自由基的能力可能是抵抗肌肉中自由基损伤的主要保护性适应措施,因此可能是抵抗少肌症发展或发作的主要保护措施,并且可能还涉及Gpx1和Gsta4。我们的发现表明,SKM和心脏中活性Nrf2的增加促进了表达抗氧化剂和抗亲电子酶基因的转录激活,这些基因在衰老过程中可能起关键作用。因此,SFN可以通过Keap1 / Nrf2途径的其他作用(例如自噬、谷胱甘肽生物合成和衰老及与衰老相关疾病中的线粒体生物发生)恢复多种细胞防御,从而有效预防心脏和SKM组织功能障碍。
总之,与年龄有关的肌病是一个内在过程,其细胞和分子发生了深刻变化,导致心脏和SKM功能受损。使用小鼠衰老模型,我们证明了SFN饮食可以恢复衰老小鼠的SKM和心脏功能。我们的研究未发现含SFN饮食对小鼠有任何不利影响。对衰老心脏中呼吸链复合物活性的保护,线粒体复合蛋白的增加以及SFN的氧化损伤降低表明SFN增强了线粒体的功能。对Gpx1和Gsta4激活通路的进一步分析值得关注,因为对相关机制的分析可能发现针对肌少症的新疗法。保护和增强Nrf2驱动的生物学功能可能是对抗老年人SKM丧失和心脏功能丧失的一种安全有效的策略。我们的发现提供了有关新型生物标志物和替代终点的机制理解和信息,可应用于临床。
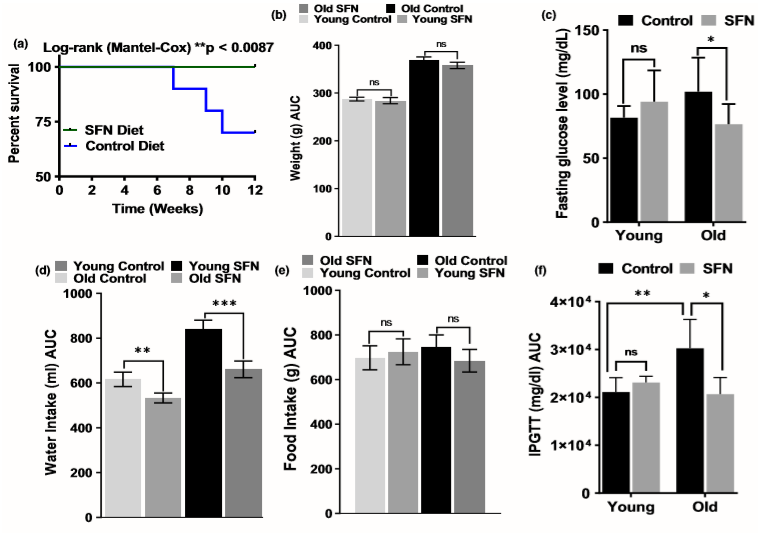
图1. 萝卜硫素(SFN)饮食对小鼠存活、体重、食物摄入、水分摄入、空腹血糖和葡萄糖耐量的影响。与对照组相比,每组补充SFN的老年小鼠的存活率均显著提高(a)(p = .0087,n = 20)。在开始SFN或对照饮食后,每周对年轻和老年小鼠的体重(b)、水摄入量(d)和食物摄入量(e)进行称重。所示数据代表平均值±SD(n = 10)。使用重复测量双向ANOVA,然后进行Bonferroni检验或未配对的Student t检验(下图),确定同一组SFN或对照饮食喂养的小鼠之间的统计学显著性。与同一组相比,* p <.05且ns =不显著。SFN还减少了空腹血糖,并改善了受葡萄糖攻击的老年小鼠的葡萄糖耐量。 (c)在禁食8小时后,测量对照或SFN饮食的年轻和老年小鼠的空腹血糖水平。显示了平均值±SD(n = 10);通过t检验,对照组或SFN日粮喂养的老年小鼠之间的差异具有统计学意义* p <.05和** p <.01。(f)禁食或SFN饮食的年轻和老年小鼠禁食4 h,并腹腔注射葡萄糖(2 g / kg体重)。通过对每只小鼠在0-120分钟之间的数值积分来计算血糖水平-时间曲线下的面积,并且显示了该面积的平均值±SD(n = 10)。年轻小鼠之间的差异无统计学意义,但使用SFN饮食的老年小鼠之间的差异明显改善。
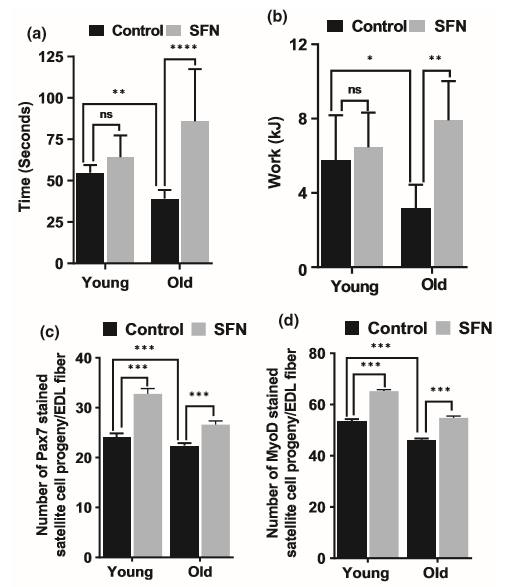
图2. 萝卜硫素(SFN)改善了老鼠的抓地力、运动表现和骨骼肌干细胞数量。(a)联合前肢抓地力测试用于测量小鼠的肌肉强度。(b)适应后,使小鼠在跑步机上以10%的斜率跑步,以增加疲劳速度。补充SFN饮食能显著改善(t检验的* p <.05和**** p <.0001)老年小鼠的运动能力和抓地力(n = 10)。将来自同一组的成批肌纤维共免疫Pax7(c)和MyoD(d)。数值代表来自平均年龄的T72肌纤维补充SFN或对照饮食的总体数据中的总体平均值。每个类别中卫星细胞的数量表示为肌纤维上存在的全部免疫染色细胞的平均百分比(通过t检验,双向方差分析,*** p <.001)。
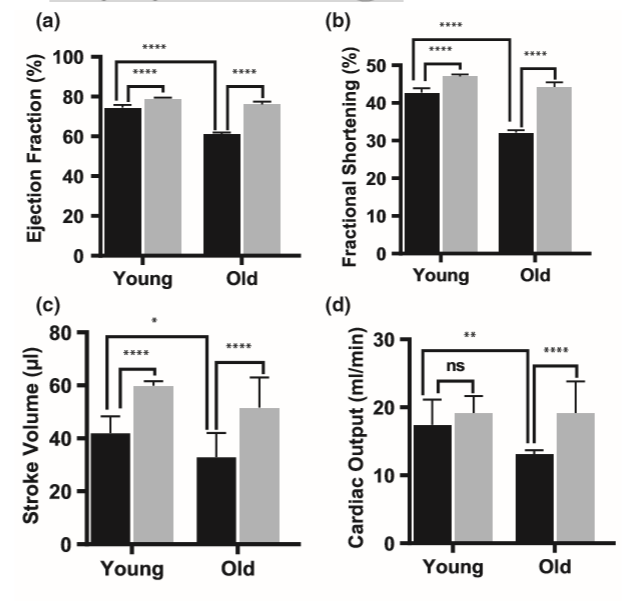
图3. 萝卜硫素(SFN)处理可保护小鼠免受年龄相关性心肌病的侵害。通过以下各项来评估心脏功能:(a)射血分数,(b)短轴缩短率,(c)每搏输出量和(d)心输出量,它们在补充了SFN饮食的老年小鼠中显著保留。黑色柱代表以对照饮食喂养的小鼠,灰色柱代表以补充SFN饮食喂养的小鼠。统计显著性*** p <.001和**** p <.0001由ANOVA和Tukey(n = 10)确定。

图4萝卜硫素(SFN)可增加ETC复杂蛋白的表达并保护衰老心脏中线粒体的超微结构。(a)SFN可以防止衰老心脏中的线粒体嵴紊乱、部分嵴溶解和基质电子密度降低(代表n = 3)。(b)NDUSF3和CORE-2的免疫组织化学检测(棕色染色)显示,SFN喂养的老小鼠左心室水平升高(代表n = 5)。(c)补充SFN可以逆转衰老心脏因氧化应激引起蛋白质硝化作用(代表n = 5)

图5. 萝卜硫素(SFN)改善了衰老心脏的ETC功能。在接受i)对照饮食或ii)SFN喂养的年轻和老年小鼠(n = 10)的新鲜心脏(a)和SKM活检组织(b)中,使用方法部分中所述的底物抑制剂滴定方法评估了复合物I,II + III的呼吸状态以及ETC的最大呼吸。根据氧气通量测量,与对照组饮食喂养的老年小鼠相比,SFN喂养的老年小鼠表现出改善的I,I + II和最大呼吸。每个条形表示平均值±SD(n = 10); 通过非配对t检验评估统计学显著性(通过t检验,ns> 0.05,** p <.01和*** p <.001)。
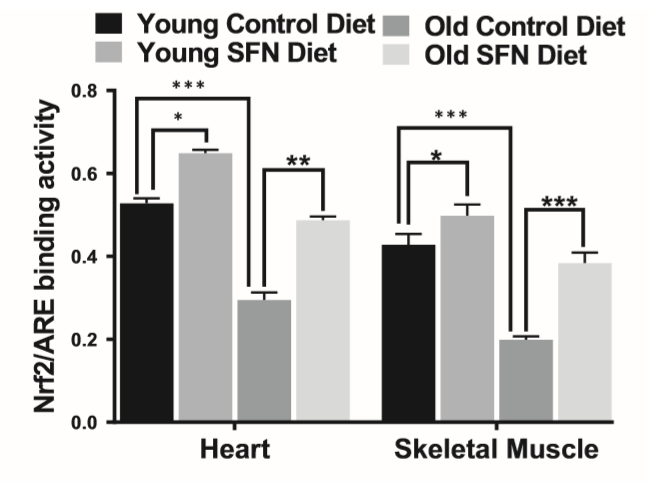
图6. 补充萝卜硫素(SFN)的饮食可改善Nrf2活性。SFN补充改善了老年小鼠心脏的Nrf2-ARE结合活性(n = 5)。统计学显著性* p <.05,** p <.01和*** p <.001是通过ANOVA和随后的Tukey多重比较检验确定的。
表1. 对照组年轻和老年小鼠以及饲喂SFN的小鼠心脏Nrf2和Nrf2靶基因转录的相对丰度。

表2. 对照组年轻和老年小鼠以及饲喂SFN小鼠骨骼肌Nrf2和Nrf2靶基因转录量的相对丰度。
年龄相关的线粒体功能障碍和氧化损伤是多种健康问题的主要原因,包括肌肉减少症和心血管疾病(CVD)。尽管Nrf2(一种调节细胞保护性基因表达的转录因子)在肌肉病变中的作用仍不清楚,但它在肌肉减少症和CVD中均显示出有益的特性。萝卜硫素(SFN)是一种激活Nrf2相关细胞保护基因的天然化合物,可对包括CVD在内的多种疾病提供保护,并且正在进行从预防癌症到降低胰岛素抵抗的各个阶段临床试验。这项研究旨在确定SFN是否可以预防与年龄相关的心脏和骨骼肌功能丧失。对2个月大和21至22个月大的小鼠进行常规啮齿动物饮食或补充SFN的饮食12周。在研究完成时,检测了骨骼肌和心脏功能、线粒体功能和Nrf2活性。我们的研究表明,与年轻组小鼠相比,老年对照组小鼠的Nrf2活性和线粒体功能显著下降,并且骨骼肌和心脏功能丧失。在老年小鼠中,SFN恢复了Nrf2活性、线粒体功能、心脏功能、运动能力、葡萄糖耐量以及骨骼肌卫星细胞的激活/分化。我们的结果表明,与年龄相关的Nrf2信号传导活性下降和与之相关的线粒体功能障碍可能与年龄相关疾病的发展有关。因此,通过SFN恢复Nrf2活性和内源性细胞保护机制可能是一种安全有效的策略,可以防止由于衰老而引起的肌肉和心脏功能障碍。
关键词:心脏功能、线粒体功能障碍、Nrf2、氧化应激、肌肉减少症、萝卜硫素
1. 前言
衰老是受内在和外在因素共同调节的复杂生物过程。衰老的机制包括活性氧(ROS)、端粒缩短和激素变化的作用。 年龄相关的线粒体功能障碍和氧化损伤是多种健康问题的主要原因,包括肌肉减少症和心血管疾病(CVD)(Miller et al.,2012)。已经评估了几种化学和功能不同的ROS和抗氧化剂产品的清除剂缓解年龄引起的肌肉丢失的能力(Barrera等人,2018),但收效甚微。它们缺乏益处的主要原因可能包括细胞系统中生物利用度低和/或对氧化剂和亲电试剂的清除能力低。因此,促进内在抗氧化剂和细胞保护途径的活性可能代表了一种对抗老年人肌肉功能丧失的更有效策略。
对于细胞适应氧化应激和亲电子应激至关重要的酶(和其他蛋白质)的转录调控是由抗氧化剂反应性DNA元件(ARE)控制的,也称为亲电反应元件(Suh等人,2004)。细胞对亲电应激反应的主要调节因子是转录因子Nrf2(核红细胞-2-p45相关因子-2),被证明对解毒基因活性至关重要(Itoh等,2004),包括在哺乳动物的心脏细胞中以及心血管系统的其他组成部分(Dastani等,2012)。尽管Nrf2在肌病中的作用尚不明确,但其下调与肌肉减少症和CVD相关(Gounder等,2012)。在生理条件下,Nrf2通过其阻遏因子,kelch样ech结合蛋白1 (Keap1)结合在细胞质中。Keap1调节亲电体对Nrf2的降解(Itoh等,2004)。亲电物,包括天然化合物萝卜硫素(SFN),通过Keap1的硫醇修饰激活抗氧化和细胞保护途径,使其在细胞质中与Nrf2分离。释放后,Nrf2积聚在细胞核中,与小的Maf蛋白异源二聚体,并通过其AREs激活靶基因的转录(Cao等人,2006)。 Nrf2靶基因编码参与细胞保护反应的蛋白质,包括细胞抗氧化剂和抗亲电子活性以及重要的细胞抗氧化剂谷胱甘肽(GSH)的生物合成。迄今为止,亲电代谢中编码酶的所有已知基因以及众多抗氧化酶均在基础表达水平受到调控,并经常被Nrf2诱导(Dinkova-Kostova&Abramov,2015)。
产生过量ROS的线粒体缺陷对细胞有害,并经常通过自噬/线粒体途径清除,然后诱导线粒体生物发生,以试图恢复线粒体的质量和功能(DinkovaKostova&Abramov,2015)。Nrf2通过直接激活保护机制来维持氧化应激期间的线粒体形态和完整性(Dinkova-Kostova&Abramov,2015)。
SFN是十字花科蔬菜中的一种植物化学物质,是一种无毒化合物,因其在不同疾病模型中的抗衰老、抗癌、抗糖尿病、抗微生物和化学预防活性而被公认(Kensler等,2013)。增强的Nrf2信号传导和随后的细胞保护性基因激活被认为是SFN作用的主要机制(Bai等人,2013)。SFN已显示出减少小胶质细胞介导的神经炎症的潜力,并可以减轻神经性行为缺陷并减轻阿尔茨海默氏病模型小鼠的αβ负担(Uddin等,2020)。SFN改善肌肉功能,改善病理学,保护营养不良的肌肉并减轻肌肉发炎(Sun等,2015)。SFN还通过Akt/Foxo信号传导抑制地塞米松诱导的肌管肌肉萎缩(Son等人,2017)。此外,SFN可以通过促进抗氧化剂机制修复与年龄有关的心血管疾病中的血管平滑肌细胞功能障碍,并防止皮肤衰老(Sedlak等人,2018)。目前,一些正在进行的临床前和临床试验研究了其对癌症、胰岛素抵抗、精神分裂症和自闭症的作用(Kensler等,2013; K. Singh等,2014)。
因为已知SFN可以激活Nrf2并减轻氧化损伤,所以在这项研究中,我们研究了SFN处理是否可以恢复老年小鼠的Nrf2活性、线粒体功能以及骨骼肌和心脏功能。我们还评估了SFN处理和对照小鼠的骨骼肌和心脏中的转录组改变。在这项研究中,我们为上调Nrf2活性作为一种缓解年龄诱导的肌肉和心脏功能障碍的新治疗策略奠定了基础。
2. 结论
2.1 SFN提高生存率,但不影响动物体重
在21-22个月大时,将雄性C57BL / 6小鼠置于含有SFN的饮食或对照饮食(每组n = 20小鼠)中,共12周。在SFN给药期间,对照组中有6只小鼠死亡。另一方面,SFN饮食组中的所有小鼠均存活到实验结束,直到它们达到24至25个月大时为止(图1a)。膳食补充SFN并不会影响年轻或成年小鼠的体重。正如预期的那样,在两个饮食组中,年轻小鼠的体重增长都快于老年小鼠(图1b)。另一方面,补充SFN会影响进水量(图1c)。与对照组饮食相比,SFN饮食喂养的年轻小鼠的饮水量显著降低。在年长的动物群中也观察到了同样的情况。但是,食物摄入量不受影响(图1d)。此外,补充SFN不会影响年轻和老年小鼠的血液学参数(表S1)。
2.2 SFN可以降低老年小鼠的空腹血糖并改善其葡萄糖耐量
在SFN饮食中,年轻小鼠的空腹血糖水平不高于年轻对照小鼠(图1e)。另一方面,饲喂SFN日粮的老年小鼠与老年对照组小鼠(102±9 mg / dl)相比,葡萄糖水平显著降低(平均±SD:77±16 mg / dl)。在两个饮食组中,葡萄糖水平都在C57BL / 6小鼠的正常范围内或以下(Surwit等,1988)。由于对照组和治疗组动物的空腹血糖水平差异不大,因此进行了腹膜内葡萄糖耐量试验。SFN处理不影响年轻动物对葡萄糖给药的反应(图1f)。但是,与对照组相比,在接受饮食控制的老年小鼠中,注射葡萄糖后血糖水平显著增加。此外,SFN治疗逆转了这种血糖升高的情况,并改善了老年动物的葡萄糖耐量(图1f)。
2.3 SFN可改善老年小鼠的运动能力
我们测试了SFN治疗是否会改善肌肉力量和运动能力。为此,我们研究了接受对照或SFN饮食喂养的年轻和老年小鼠,进行了不随意的体育锻炼来测试其肌肉力量。首先,我们通过将它们固定在粗线上来测试它们的抓地力。用对照饮食喂养的年轻和老年小鼠能够分别平均维持54±5和39±5 s。用SFN饮食的年轻和老年小鼠能够保持更长的时间,分别为64±13和86±12 s(图2a)。令人惊讶的是,喂SFN的老年小鼠比喂SFN的年轻小鼠能坚持更长的时间。此外,我们比较了对照组和SFN处理的年轻小鼠和老年小鼠的运动能力,方法是让它们在电动,速度可控的模块化跑步机系统上运行(Handschin等,2007)。与年轻的同龄小鼠相比,接受对照饮食的老年小鼠的运动能力较低(图2b)。饲喂富含SFN的饮食可以显著改善老年小鼠的运动能力。由SFN喂食的老年老鼠在跑步机上的表??现与年轻动物相似。
2.4 SFN增加了骨骼肌干细胞的数量及其功能
从下肢的EDL肌肉分离的肌纤维培养72小时。固定了卫星细胞衍生的成肌细胞,并对Pax7和MyoD进行了免疫染色,以此作为卫星细胞增殖和分化的指标(图S1-S8)。大部分卫星细胞表达Pax7,甚至更多表达MyoD(图2c)。Pax7在SKM干细胞中的表达不影响MyoD表达,并有助于SKM的生长和再生(Zammit等,2006)。 在SFN饮食的年轻和老年小鼠中,Pax7-(图2c)和MyoD(图2d)阳性卫星细胞子代的比例显著高于其年龄匹配的对照组。SFN治疗可能会增加新的卫星细胞形成,从而满足肌肉稳态的常规需求,或者可能是偶发性的肥大或修复需求。
2.5 SFN减少了SKM老化,氧化和凋亡的标志
组织学显示,使用SFN饮食2个月后,载玻片的横截面积更多地被肌纤维占据(图S9)。同样,肌肉肌抑制素(SKM质量的负调节剂)的染色也较少(图S10)。最后,我们发现有证据表明SFN可以降低8OHdG(一种氧化的标志物)(图S11)和Tunel染色(一种凋亡的替代标志物)(图S12)。
2.6 SFN治疗老年小鼠也可改善心脏功能
已知心脏/呼吸功能下降会限制老年人的运动能力(Farkhooy et al。,2018)。心脏衰老是一个内在的过程,具有深刻的细胞和分子变化,可导致心脏功能受损(Vigorito&Giallauria,2014)。我们检查了SFN是否能恢复老年小鼠的心脏功能。与对照饮食的年轻小鼠相比,对照饮食的老小鼠射血分数(61.0±1.0%)、短轴缩短率(32.1±0.7%)和每搏输出量(32.8±9.2 µl)均降低(图3a-c)。另一方面,补充SFN可以改善老年小鼠的射血分数(76.0±1.4),短轴缩短率(44.2±1.3%)和每搏输出量(51.6±11.3 µl)。尽管SFN还可以显著增加年轻小鼠的这些参数,但对老年小鼠的射血分数和分数缩短的影响更大。补充SFN对年轻小鼠的心输出量无明显影响,而老年小鼠的心输出量则有明显改善(图3d)。结果,SFN喂养的老年小鼠的心输出量与年轻对照组的相似。我们得出的结论是,SFN喂养的老年小鼠对与年龄相关的心脏功能丧失具有抵抗力。
2.7 SFN保护衰老心脏中的线粒体超微结构
线粒体是高能量需求型心肌细胞的主要能源,占据其细胞体积的40%以上(约5,000个线粒体/心肌细胞)。心肌细胞功能严重依赖于这些细胞器的健康状况(Strom等,2016)。此外,与年龄相关的线粒体功能障碍可能是由于氧化损伤导致线粒体超微结构的变化所致(Vays等,2014)。为了确定SFN是否能预防与年龄相关的心脏线粒体超微结构变化,我们使用透射电子显微镜比较了对照组和SFN喂养的老年小鼠心脏之间的线粒体形态。结果表明,SFN保护线粒体免于与年龄相关的嵴紊乱、部分嵴溶解和基质电子密度降低的影响(图4a)。由于参与氧化磷酸化的酶位于线粒体内膜上,嵴的表面积和数量通常与细胞的代谢活性相关(Arismendi-Morillo,2011)。为了进一步验证在用SFN处理的老年小鼠心脏中观察到的电子转运链(ETC)复合物活性的增加/保护,我们用针对复合物I(NDUFS3-a核DNA编码亚基)和复合物III的蛋白质的抗体进行了免疫染色(CORE2-线粒体编码的亚基)。与年龄匹配的未治疗动物的心脏相比,SFN处理后衰老的心脏中的线粒体复合物I和III亚基蛋白表达增加(图4b)。这些数据表明,SFN可能维持线粒体功能的一种机制是诱导线粒体蛋白表达。另外,在老年小鼠的心肌线粒体中发现的蛋白质表达下降可能是由于氧化损伤。左心室组织的免疫组织化学分析显示,SFN喂养的老年小鼠具有较低水平的硝基酪氨酸蛋白加合物(氧化应激的标志物)(图4c)。这些作用可能有助于增加老年小鼠心脏的自噬,并通过SFN减轻自噬,如LC3-II在老年小鼠中的出现及其在SFN治疗后的消失所证明的(图S13)。
2.8 SFN改善老年小鼠的线粒体功能
心脏组织的线粒体特别丰富,可以满足其高代谢需求。线粒体呼吸复合物的破坏可导致氧化剂的产生,ATP耗竭和心肌细胞功能障碍。线粒体功能一直被认为在衰老过程中会下降,从而导致氧化应激增加(Hebert等,2015)。我们通过高分辨率呼??吸测定法测量了心脏和SKM组织的线粒体功能。与年轻的小鼠心脏相比,老年人的ETC活性低约30%,SFN保护ETC免受这种与年龄相关的氧通量下降(图5a)。此外,在SFN喂养的动物中ETC复合物I,I + II和最大呼吸的活性增加,但这种增加仅在老年小鼠中才显著。SFN喂养动物的SKM的线粒体中也发现了这种类似趋势,但并未达到显著水平(图5b)。
2.9 SFN恢复老年小鼠心脏和SKM中的Nrf2活性
除了对线粒体的影响外,Nrf2功能与年龄相关的下降已有充分的文献记载(Gounder等,2012),但尚不清楚SFN是否能在老年人群中恢复Nrf2的活性。最近,我们报道Nrf2活性随SFN处理而增加,并且是保护心脏免受诸如阿霉素和电离辐射之类的氧化剂伤害的关键(Boerma等,2015; P。Singh等,2015)。因此,我们比较了Nrf2 ARE结合活性在年轻和老年小鼠的心脏和SKM中的作用。与年龄匹配的对照组相比,SFN喂养的年轻和老年小鼠的心脏和SKM的核提取物中Nrf2活性均上调(图6)。
2.10心脏和骨骼肌的qRT-PCR结果进一步描述了SFN发挥作用的潜在机制
为了进一步阐明SFN介导的小鼠心脏和SKM功能衰老增强的机制,我们检查了许多与氧化剂和亲电子代谢相关的基因的表达。选定的基因参与抗氧化剂,抗亲电子活性,哺乳动物的寿命以及谷胱甘肽的合成和损失(表1和2),所有这些基因均受Nrf2的调控(Hayes&Dinkova-Kostova,2014)。我们检查了过氧化氢酶,Sod1,Sod2,HO1,Pxdn,Gpx1,HO-1,Gsta4,Akr3,Akr7,Akr 8,Sirt1,Pgc1,Gclc,Gclm和Nrf2的心脏和SKM转录水平(全表见表1基因)。在对Gclc,Gclm,Gsta4,Nqo-1,Ho-1和Sod2启动子区域进行的计算机或体外分析中,发现了Nrf2的共有ARE结合位点(Tonelli et al。,2018)。
与饲喂对照饮食的小鼠相比,SFN饮食喂养的年轻和老年小鼠心脏中的许多抗氧化剂和抗亲电子基因的心脏转录水平显著增加(表1)。新陈代谢的这种变化可以解释为什么含有SFN的饮食具有心脏保护作用,并能增强运动能力。在对照组中,大多数检查的基因以较低的水平表达。与SFN喂养的动物相比,老年对照动物产生的较低水平的抗氧化剂和抗电子防御机制可能有助于心脏的氧化损伤。在用SFN日粮喂养的动物中,某些转录本的水平,特别是Sod1,Sod2,Cat和Nrf2,在老小鼠的心脏中降低,并且在日粮中被SFN基本上恢复了。Akr3证明了老年小鼠心脏的增加,并且在饮食中通过SFN得以部分恢复。
表2给出了来自相同小鼠的SKM的类似实验的结果。在这种情况下,这些基因中的更多基因在老年小鼠中被下调,并且在饮食中被SFN很大程度上恢复:Cat,Sod1,Gpx1,Gsta4和Nrf2。尽管Sod2,Pxdn,Gclc和Gclm不受衰老的影响,但在老年小鼠的饮食中,它们也被SFN上调。总之,这些结果表明Nrf2途径是在SFN治疗后在老年小鼠中诱导的,并增强了SKM中的保护机制,可能甚至超过心脏。总而言之,这些结果表明Nrf2途径是在SFN处理后在年轻和老年小鼠中诱导的,并增强了保护机制。
3. 讨论
SFN可以延长老年小鼠的寿命,降低其患糖尿病的可能性,并提高其运动能力。运动能力增加的部分原因当然是由于SKM功能增强,但某些原因可能是由于保留了心脏功能。此外,在研究期间比较了使用SFN或对照饮食的成年小鼠的存活曲线,显示了显著的保护作用,从采用SFN饮食的老年动物的死亡率中得出结论(对数秩(Mantel-Cox)检验,p <.0087; Gehan–Breslow–Wilcoxon检验,p <.0089)。
我们使用年轻和老年的C57BL / B6J小鼠进行了SFN饮食研究,因为该小鼠寿命相对较长,并且死因各异,但肿瘤发生率低(Treuting等,2008)。先前的研究表明,SFN治疗会降低以高脂饮食或以高脂高蔗糖饮食喂养的肥胖小鼠为食的小鼠的体重增加和食物摄入,并改变其代谢参数(Shawky等人,2016; Shawky&Segar, 2018)。使用更正常的饮食,我们发现SFN对体重或食物摄入量没有影响,尽管我们确实发现用SFN喂养的老年小鼠的水摄入量减少了。我们推测增加的肌肉质量可能弥补了水摄入减少所引起的体重差异。小鼠对血液学参数无统计学显著影响(表S1)。使用SFN饮食的老年小鼠的空腹血糖水平比老年对照动物的空腹血糖水平稍低,但仍保持在正常范围内。如图1f所示,IPGTT在年轻小鼠中是正常的,但是与SFN饮食中的老年小鼠相比,对照饮食中的老年小鼠中胰岛素抵抗变得明显。个别年龄较大的小鼠(对照组和SFN饮食)对胰岛素的反应之间的差异表明,采用对照饮食的老年小鼠开始出现代谢综合征和病理。这些结果表明,SFN饮食可能对这些老年小鼠的胰岛素抵抗具有有益的作用。
目前的研究结果包括,在使用SFN饮食的老年小鼠中,跑步机的运动能力和对前肢握力的反应明显改善。饲喂SFN日粮的老年小鼠的跑步机运动能力大约是饲喂对照饮食的小鼠的1.5倍。当比较前肢的握力时,用SFN喂养的老年小鼠挂在电线上的时间是使用对照饮食的老年小鼠的两倍以上。用SFN喂养的老年小鼠的表现甚至比年轻的小鼠还要好。用SFN喂养的小鼠的EDL肌纤维中Pax7和MyoD阳性卫星细胞的子代增加,表明老年小鼠的肌肉再生得到改善。总体而言,我们已经证明SFN饮食干预可以改善老年小鼠的运动表现,抓地力和增加肌肉干细胞的形成,这表明SFN可能是减少衰老人群肌肉减少症的一种新型治疗方法。
SFN还对老年小鼠的心脏功能产生治疗作用。我们的结果表明,SFN治疗可显著改善老年小鼠的心脏射血分数,缩短分数,中风量和心输出量,使老年小鼠的静息心功能达到年轻小鼠的水平。这些特定的小鼠似乎比大多数其他研究的小鼠品系更容易发生收缩功能障碍,这些品系主要患有舒张功能障碍(de Lucia等人,2019)。然而,有报道说衰老小鼠的左心室射血分数降低导致收缩功能障碍(Han等,2020; Quarles等,2020),并且我们以前已经证明SFN可以防止阿霉素诱导的氧化应激左心室射血分数和分数缩短的异常(Benes et al。,2013; Bose et al。,2018; Singh et al。,2015)。
这项研究有几个重要的局限性。它仅在雄性小鼠中进行;因此,性别差异是未知的。此外,这项研究是在相对较小的样本量下进行的,仅使用了两个年龄组,并且持续时间相对较短(12周)。我们尚未完全描述SKM与心脏受SFN影响的机制的差异。最后,将结果转换为人类是未知的,尤其是在心力衰竭方面。然而,很明显,老年人的骨骼肌和心肌功能均下降,并且极大地降低了生活质量。在这种情况下,SFN可能是一种有希望的策略,可以减轻这些组织的衰老过程。
衰老心脏中ROS的产生增加可导致心肌细胞的坏死和凋亡以及成纤维细胞的增殖以及胶原蛋白的过量生成和纤维化的发展,以及线粒体的损??害;这些事件随后导致心脏重塑和功能障碍(Dai等,2014)。不幸的是,心脏中的抗氧化剂和亲电子保护性酶水平较低,使其特别容易受到自由基的损害(Singh等,2015)。年龄相关的线粒体功能障碍是衰老的重要部分。这些能量源能够自我复制,但随着年龄的增长,会通过各种机制(包括氧化/亲电子应力的持续恶性循环)而失效。为了进一步确定SFN在衰老过程中在线粒体功能和形态中的作用,我们评估了线粒体的超微结构,关键的电子转运链复合物蛋白表达以及小鼠心脏中氧化应激的标志物以及心脏和SKM中的线粒体呼吸复合物。氧化剂和亲电子试剂的增加,导致衰老期间的Nrf2活性和Nrf2响应基因,修饰了参与能量代谢的几种线粒体蛋白,并导致嵴的混乱。从我们对心脏和SKM活组织检查的高分辨率呼吸测定研究中获得的数据表明,衰老会损害复杂的I,I&II和最大呼吸功能,但这种影响在心脏中更为明显。由SFN喂养的年老小鼠的线粒体复杂活动的心脏已完全恢复,接近年轻对照组的水平。由于这些活动在老年动物的心脏活检中严重降低,因此恢复具有重要意义。喂食老年动物SFN后,CORE-2和NDUFS3表达增加,线粒体嵴混乱、部分嵴溶解、电子透明基质以及硝基酪氨酸蛋白加合物的水平降低,说明也可能有助于改善线粒体功能以及通过Nrf2依赖性细胞防御激活的SFN保护作用。我们认为通过SFN药理激活Nrf2可能会通过增强2相排毒和抗氧化酶活性并保留电子传输链(ETC)复杂功能来消除衰老过程中产生的过量ROS。由于在线粒体较少的SKM中,其对线粒体功能的影响不太明显,因此SFN可能对SKM发挥其他额外作用。
细胞中的累积氧化损伤是细胞衰老和衰老的基本特征。与年龄相关的氧化应激会促进心肌纤维化,左心室硬化的进行性增加以及心脏功能障碍,并且可能是导致肌肉无力和肌肉减少症的因素之一。与年龄相关的Nrf2功能障碍可降低抗氧化剂和抗亲电子酶的活性以及线粒体功能,导致体内稳态失调,并损害细胞的修复和再生能力。我们以前已经在体外和体内证明了SFN保护和增强Nrf2信号传导,通过激活细胞保护性基因以抵抗辐射和阿霉素引起的心脏毒性以及其他相关疾病(Chapple等,2012)。我们最近证明,SFN增强了小鼠心脏中的Nrf2信号传导和细胞保护性基因活性,从而保护免受氧化损伤(Singh等人,2015)。
在本研究中,我们研究了饮食中的SFN是否能激活两个最重要的功能组织SKM和心脏中的Nrf2活性和响应基因。我们确定了喂食SFN或对照饮食的年轻和老年小鼠的SKM和心脏中活性核Nrf2的量。SKM和SFN日粮喂养的年轻和老年小鼠的心脏显示ARE结合活性显著增加,这与对照饮食中老年小鼠中观察到的活性显著降低相反。老年对照组小鼠中Nrf2活性的下降清楚地表明,SKM和心脏(已经表达了低水平的保护酶)无法保护自己免受氧化和亲电攻击并正常运行。因此,在这项研究中,我们首次证明了衰老的心脏和SKM中氧化应激的增加与Nrf2调节异常引起的保护性反应失败有关,并且可以通过让老年小鼠(21到22个月大的小鼠)服用SFN来减轻这一过程。
检测了对照组和SFN饲料喂养小鼠心脏(A)和SKM(B)中负责表达抗氧化、抗亲电性和谷胱甘肽合成途径酶的基因转录水平。(A)在心脏中,过氧化氢酶、Sod1、Sod2和Nrf2与衰老小鼠的功能降低以及SFN的恢复表现出最佳的相关性。此外,Akr3在老年小鼠中增加,SFN减少了这种增加。Sod1和Sod2保护线粒体基质中的线粒体复合物和活性酶,并维持体内稳态和线粒体完整性(Piao等人,2010)。(B)在SKM中,可能的因素众多:过氧化氢酶、Sod1、Gpx1、Gsta4和Nrf2。心脏和SKM中受影响的基因之间只有部分重叠并不奇怪,特别是考虑到衰老和SFN对SKM中线粒体的影响较小。过氧化氢酶和Sod1可能在线粒体水平上发挥某些作用,但也可以增加抗氧化剂和抗电子防御机制的水平,并有助于降低脂质过氧化产物和4-HNE的浓度,从而恢复老年小鼠SKM的功能。酶促清除超氧化物自由基的能力可能是抵抗肌肉中自由基损伤的主要保护性适应措施,因此可能是抵抗少肌症发展或发作的主要保护措施,并且可能还涉及Gpx1和Gsta4。我们的发现表明,SKM和心脏中活性Nrf2的增加促进了表达抗氧化剂和抗亲电子酶基因的转录激活,这些基因在衰老过程中可能起关键作用。因此,SFN可以通过Keap1 / Nrf2途径的其他作用(例如自噬、谷胱甘肽生物合成和衰老及与衰老相关疾病中的线粒体生物发生)恢复多种细胞防御,从而有效预防心脏和SKM组织功能障碍。
总之,与年龄有关的肌病是一个内在过程,其细胞和分子发生了深刻变化,导致心脏和SKM功能受损。使用小鼠衰老模型,我们证明了SFN饮食可以恢复衰老小鼠的SKM和心脏功能。我们的研究未发现含SFN饮食对小鼠有任何不利影响。对衰老心脏中呼吸链复合物活性的保护,线粒体复合蛋白的增加以及SFN的氧化损伤降低表明SFN增强了线粒体的功能。对Gpx1和Gsta4激活通路的进一步分析值得关注,因为对相关机制的分析可能发现针对肌少症的新疗法。保护和增强Nrf2驱动的生物学功能可能是对抗老年人SKM丧失和心脏功能丧失的一种安全有效的策略。我们的发现提供了有关新型生物标志物和替代终点的机制理解和信息,可应用于临床。
图1. 萝卜硫素(SFN)饮食对小鼠存活、体重、食物摄入、水分摄入、空腹血糖和葡萄糖耐量的影响。与对照组相比,每组补充SFN的老年小鼠的存活率均显著提高(a)(p = .0087,n = 20)。在开始SFN或对照饮食后,每周对年轻和老年小鼠的体重(b)、水摄入量(d)和食物摄入量(e)进行称重。所示数据代表平均值±SD(n = 10)。使用重复测量双向ANOVA,然后进行Bonferroni检验或未配对的Student t检验(下图),确定同一组SFN或对照饮食喂养的小鼠之间的统计学显著性。与同一组相比,* p <.05且ns =不显著。SFN还减少了空腹血糖,并改善了受葡萄糖攻击的老年小鼠的葡萄糖耐量。 (c)在禁食8小时后,测量对照或SFN饮食的年轻和老年小鼠的空腹血糖水平。显示了平均值±SD(n = 10);通过t检验,对照组或SFN日粮喂养的老年小鼠之间的差异具有统计学意义* p <.05和** p <.01。(f)禁食或SFN饮食的年轻和老年小鼠禁食4 h,并腹腔注射葡萄糖(2 g / kg体重)。通过对每只小鼠在0-120分钟之间的数值积分来计算血糖水平-时间曲线下的面积,并且显示了该面积的平均值±SD(n = 10)。年轻小鼠之间的差异无统计学意义,但使用SFN饮食的老年小鼠之间的差异明显改善。
图2. 萝卜硫素(SFN)改善了老鼠的抓地力、运动表现和骨骼肌干细胞数量。(a)联合前肢抓地力测试用于测量小鼠的肌肉强度。(b)适应后,使小鼠在跑步机上以10%的斜率跑步,以增加疲劳速度。补充SFN饮食能显著改善(t检验的* p <.05和**** p <.0001)老年小鼠的运动能力和抓地力(n = 10)。将来自同一组的成批肌纤维共免疫Pax7(c)和MyoD(d)。数值代表来自平均年龄的T72肌纤维补充SFN或对照饮食的总体数据中的总体平均值。每个类别中卫星细胞的数量表示为肌纤维上存在的全部免疫染色细胞的平均百分比(通过t检验,双向方差分析,*** p <.001)。
图3. 萝卜硫素(SFN)处理可保护小鼠免受年龄相关性心肌病的侵害。通过以下各项来评估心脏功能:(a)射血分数,(b)短轴缩短率,(c)每搏输出量和(d)心输出量,它们在补充了SFN饮食的老年小鼠中显著保留。黑色柱代表以对照饮食喂养的小鼠,灰色柱代表以补充SFN饮食喂养的小鼠。统计显著性*** p <.001和**** p <.0001由ANOVA和Tukey(n = 10)确定。
图4萝卜硫素(SFN)可增加ETC复杂蛋白的表达并保护衰老心脏中线粒体的超微结构。(a)SFN可以防止衰老心脏中的线粒体嵴紊乱、部分嵴溶解和基质电子密度降低(代表n = 3)。(b)NDUSF3和CORE-2的免疫组织化学检测(棕色染色)显示,SFN喂养的老小鼠左心室水平升高(代表n = 5)。(c)补充SFN可以逆转衰老心脏因氧化应激引起蛋白质硝化作用(代表n = 5)
图5. 萝卜硫素(SFN)改善了衰老心脏的ETC功能。在接受i)对照饮食或ii)SFN喂养的年轻和老年小鼠(n = 10)的新鲜心脏(a)和SKM活检组织(b)中,使用方法部分中所述的底物抑制剂滴定方法评估了复合物I,II + III的呼吸状态以及ETC的最大呼吸。根据氧气通量测量,与对照组饮食喂养的老年小鼠相比,SFN喂养的老年小鼠表现出改善的I,I + II和最大呼吸。每个条形表示平均值±SD(n = 10); 通过非配对t检验评估统计学显著性(通过t检验,ns> 0.05,** p <.01和*** p <.001)。
图6. 补充萝卜硫素(SFN)的饮食可改善Nrf2活性。SFN补充改善了老年小鼠心脏的Nrf2-ARE结合活性(n = 5)。统计学显著性* p <.05,** p <.01和*** p <.001是通过ANOVA和随后的Tukey多重比较检验确定的。
表1. 对照组年轻和老年小鼠以及饲喂SFN的小鼠心脏Nrf2和Nrf2靶基因转录的相对丰度。
表2. 对照组年轻和老年小鼠以及饲喂SFN小鼠骨骼肌Nrf2和Nrf2靶基因转录量的相对丰度。
本文由福山生物整理翻译,转载请注明出处。