






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
Nrf2缺乏对老年骨骼肌线粒体氧化应激的影响
发表于:2021-07-06 作者:admin 来源:本站 点击量:11709
原文:Yu K , Tamura Y , Takahashi K , et al. Effects of Nrf2 deficiency on mitochondrial oxidative stress in aged skeletal muscle[J]. Physiological Reports, 2019, 7(3).
翻译:
2日本东京日本体育科学大学运动生理学系
3日本东京大学体育科学系
4日本筑波大学综合人类科学研究科
关键词 衰老,线粒体,氧化应激,骨骼肌肉
摘要 氧化应激和线粒体功能障碍与衰老过程有关。然而,核因子红细胞系2相关因子2(Nrf2)在衰老过程中对骨骼肌的作用仍有待阐明。在目前的研究中,我们评估了缺乏Nrf2,它被称为氧化还原稳态的主要调节因子,是否会促进骨骼肌中与年龄相关的线粒体功能障碍和肌肉萎缩。在这里,我们揭示了在老年Nrf2敲除的(KO)小鼠中,由于Nrf2-靶向抗氧化基因的表达降低,线粒体中氧化应激的标志物4-羟基壬烯醛和蛋白质羰基强烈升高。线粒体呼吸随着衰老而下降,然而,敲除Nrf2 和年龄匹配的野生型小鼠之间没有差异。类似地,与年轻野生型小鼠相比,老年野生型和敲除Nrf2 小鼠中的细胞色素c氧化酶活性较低。老年敲除Nrf2 小鼠肌肉中Mfn1和Mfn2 mRNA的表达较低。在老化的敲除Nrf2小鼠中,每消耗氧气的线粒体活性氧产生增加。敲除Nrf2 对体重归一化的肌肉质量没有影响。这些结果表明,Nrf2缺乏会加剧与年龄相关的线粒体氧化应激,但不会影响骨骼肌呼吸功能的下降。
引言
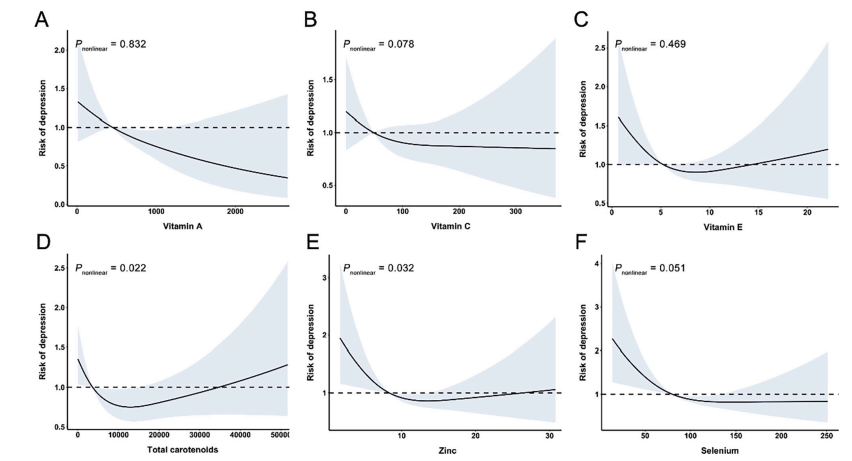
衰老的特征在于骨骼肌质量和功能的丧失,这种现象称为肌肉减少症。这种肌肉功能的丧失导致身体虚弱和死亡的风险增加(Cruz-Jentoft等,2010)。因此,了解肌肉萎缩随老化的机制对于识别治疗目标和促进健康非常重要。衰老过程被认为是由氧化应激驱动的,最初由Harman 于1956年提出作为自由基理论。鉴于线粒体电子传递链是活性氧(ROS)的主要来源,线粒体被认为在肌肉减少症中起着至关重要的作用。与该假设一致,许多研究表明老年骨骼肌中ROS产生增加,与线粒体功能障碍和线粒体细胞凋亡易感性增加相关(Chabi等2008; Dai等2014)。然而,升高的ROS产生是否与肌肉萎缩和衰老有因果关系仍待澄清。
Nrf2是一种转录因子,被认为是抗氧化基因的关键调节因子(Motohashi和Yamamoto 2004)。响应于氧化应激,Nrf2易位到细胞核中并与其靶抗氧化基因的抗氧化反应元件结合。我们以前曾报道Nrf2缺乏会加重年轻小鼠骨骼肌中去神经支配诱导的氧化应激,而对肌肉质量的损失几乎没有影响(Kitaoka等,2016)。然而,据报道,Nrf2缺乏对抗氧化酶的影响在老年骨骼肌中比在幼小动物肌肉中更大(Miller等,2012)。有趣的是,Nrf2的消融导致与年龄相关的氧化应激状态中的肌肉再生受损(Narasimhan等,2014)。为了探讨Nrf2信号可能参与衰老过程,我们评估了Nrf2的缺乏是否促进骨骼肌中与年龄相关的线粒体氧化应激和肌肉萎缩。
方法
动物和实验设计
敲除Nrf2小鼠获自Jackson 实验室(Bar Harbor,ME)。如先前报道的那样通过尾部DNA的PCR分析对小鼠进行基因分型(Kitaoka等,2016)。将动物分组饲养(每笼3-4只),在空调房间中,在12小时光照,12小时黑暗环境中循环,随意给予标准饲料和水。使用老年雄性(22个月大)Nrf2敲除小鼠和年轻(4个月大)和年龄匹配(22个月大)雄性野生型C57BL / 6J(WT)小鼠(每组n = 6-7)。通过颈脱位使动物安乐死,迅速取出肌肉,快速冷冻,并在-80℃下储存。所有实验均由东京大学动物实验委员会批准。
RNA分离和实时定量PCR
在Trizol试剂(Life Technologies,Gaithersburg,MD)中将腓肠肌在冰上匀浆,然后用氯仿分离成有机相和水相。在用乙醇沉淀后,使用RNeasy Mini试剂盒(Qiagen)从水相中分离总RNA。在通过分光光度法(Nanodrop ND1000,Thermo Scientific,Waltham,MA)测量RNA浓度后,使用高容量cDNA逆转录试剂盒(Applied Biosystems,Foster City,CA)进行第一链cDNA合成。Nrf2,Nqo1(NAD(P)H醌氧化还原酶1),Cat(过氧化氢酶),Gclc(谷氨酸-半胱氨酸连接酶催化亚基),Sod1(超氧化物歧化酶1),Sod2,Fis1(裂变,线粒体1),Drp1的表达(启动相关蛋白1),Mfn1(mitofusin 1),Mfn2,Opa1(视神经萎缩1)使用Thermal Cycler Dice Real-Time System和SYBR Premix Ex taq II(Takara Bio,Shiga,Japan)定量。所有样品一式两份进行。Tbp(TATA盒结合蛋白)用作对照基因,其表达在各组之间没有改变。上述基因的正向和反向引物如表1所示。

线粒体分离
如前所述使用差速离心分离线粒体级分(Tamura等,2015)。简言之,将股四头肌肌肉在线粒体分离缓冲液(67mm蔗糖,50mm Tris,50mm KCl,10mm EDTA和0.2%(w / v)无脂肪酸的牛血清白蛋白,pH7.4)中轻度均质化。将匀浆在4℃下以700g离心15分钟,并将上清液在12,000g下离心20分钟。洗涤沉淀,并重悬于线粒体分离缓冲液中。在分离程序后,使用二辛可宁酸(BCA)蛋白质测定法(Pierce,Rockford,IL)定量样品的总蛋白质含量。线粒体样品用于分析4-HNE,蛋白质羰基,线粒体呼吸和ROS产生。我们通过蛋白质印迹使用针对甘油醛-3-磷酸脱氢酶(细胞溶质标记物)和细胞色素c氧化酶(COX)亚基IV(线粒体标记物;数据未显示)的抗体证实了线粒体部分的纯度。此外,通过在单独的实验中添加外源细胞色素来验证我们的线粒体分离方法的完整性。
整个肌肉裂解物
腓肠肌在放射免疫沉淀试验(RIPA)缓冲液(25 mmol / L Tris HCl,pH 7.6,150 mmol / L NaCl,1%NP-40,1%脱氧胆酸钠和0.1%十二烷基硫酸钠[SDS])中均质化,用蛋白酶抑制剂物(Complete Mini,ETDA-free,Roche Applied Science,Indianapolis,IN)和磷酸酶抑制剂(PhosSTOP,Roche Applied Science)混合。使用BCA蛋白质测定法(Pierce)定量样品的总蛋白质含量。
蛋白质印迹
将等量的蛋白质加载到10%SDS-PAGE凝胶上并通过电泳分离。 将蛋白质转移到聚偏二氟乙烯(PVDF)膜上,蛋白质印迹使用4-HNE(4-羟基壬烯醛; ab48506) ,Total OXPHOS Rodent WB Antibody Cocktail (ab110413),Fis1(ab96764),Drp1(ab56788),Mfn2(ab 124773) 来自美国剑桥;Opa1(#612606)来自日本东京。胭脂红染色用于验证一致负荷。使用C印迹扫描仪(LI-COR,Lincoln,NE)扫描并定量印迹。
蛋白质羰基含量
用市售试剂盒(#ROIK03; SHIMA 实验室,东京,日本)测量蛋白质羰基含量。在如上所述将线粒体蛋白转移至PVDF膜后,使膜与二硝基苯肼(DNPH)反应,然后进行Western印迹。
酶活性
将胫骨前肌在100(v / w)的100mmol / L磷酸钾缓冲液中匀浆。按照已建立的方案(Spinazzi等,2012),通过分光光度法测量柠檬酸合酶(CS)和COX的最大活性。如前所述(Hadwan,2018)使用分光光度法测定过氧化氢酶活性。使用Superoxide Dismutase Assay Kit(706002,Cayman,Ann Arbor,MI)按照制造商的说明测定总SOD(Mn-SOD和Cu / Zn-SOD)活性。
线粒体呼吸
将新鲜分离的线粒体(60μg)在反应缓冲液(250mmol / L蔗糖,10mmol / L Tris,1mmol / L MgCl 2)中温育。 使用具有MitoXpress Xtra荧光传感器试剂(Agilent Technology,Santa Clara,CA)的Tecan Spark多模板读数器测量线粒体氧消耗,以测量溶解氧水平(Ex:380nm / Em:670nm)。通过添加10mmol / L琥珀酸盐和1mol / L鱼藤酮和2.5mmol / L ADP来刺激复合物II驱动的III期呼吸。使用操作软件计算每分钟的相对荧光变化。
线粒体活性氧生成
将新鲜分离的线粒体(20μg)在线粒体呼吸缓冲液和50μl/ L 2'7'-二氯荧光素(DCF)中孵育。通过添加10mmol / L琥珀酸盐和11mol / L鱼藤酮和2.5mmol / L ADP,在状态III呼吸条件下测量ROS发射。使用Tecan多模板读数器测量相对荧光变化(Ex:480nm / Em:520nm)。
统计分析
数据表示为平均值±标准误差(SEM)。进行单向方差分析(ANOVA),然后进行Bonferroni多重比较试验(GraphPad Prism 6.0,La Jolla,CA)。统计学显着性定义为P <0.05。
结果
骨骼肌质量和肌肉减少指数
动物特征如图1所示。老年敲除Nrf2小鼠体重比老年野生型小鼠轻(图1A)。为了研究衰老和Nrf2缺乏对骨骼肌质量的影响,我们测量了腓肠肌和胫骨前肌的绝对质量,以及肌肉减少指数(肌肉质量/体重)。与老年野生型小鼠相比,老年敲除Nrf2小鼠的绝对肌肉质量较低(图1B)。衰老降低肌肉减少指数,然而,Nrf2缺乏没有影响(图1C)。
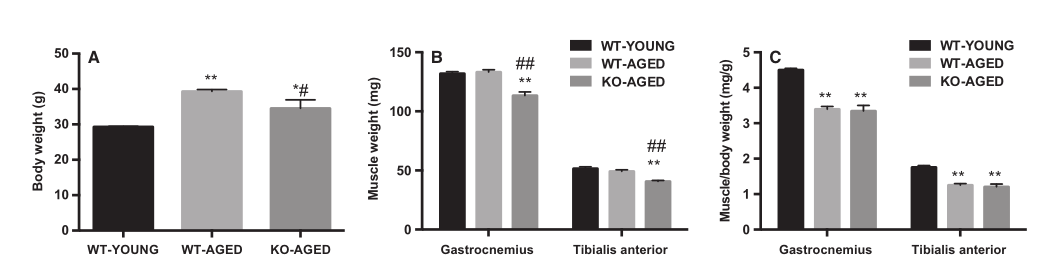
图1 老年敲除Nrf2小鼠的体重,骨骼肌质量和肌肉减少指数。 (A)体重。(B)腓肠肌和胫骨前肌肉质量。 (C)肌肉减少指数(每体重肌肉质量)。 数据表示为平均值±SEM。每组n = 6-7。* P <0.05 ** P <0.01,与年轻野生型有显着差异。#P <0.05 ## P <0.01,与老年野生型相比有显着差异。WT,野生型; KO,敲除。
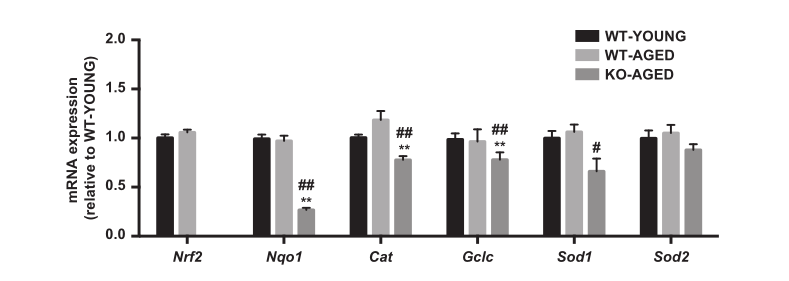
图2.老年敲除Nrf2小鼠中的Nrf2靶抗氧化基因。 数据表示为平均值±每组SEM n = 5-7。* P <0.05 ** P <0.01,与年轻野生型有显着差异。#P <0.05 ## P <0.01,与老年野生型的差异。 WT,野生型;KO,敲除。
抗氧化基因表达和氧化应激
与野生型小鼠相比,通过不可检测的mRNA表达证实Nrf2的敲除。衰老对Nrf2及其主要靶基因(Nqo1,Cat,Gclc,Sod1和Sod2)的mRNA表达没有影响。Nrf2靶向抗氧化基因的表达在老年敲除Nrf2肌肉中减少,除Sod2,其他没有改变(图2)。为了检测线粒体氧化损伤,我们测量了线粒体组分中4-HNE(脂质过氧化标记物)和蛋白质羰基含量(蛋白质氧化标记物)的水平。敲除Nrf2小鼠显示4-HNE(图3A)和蛋白质羰基含量显着增加(图3B)。与年轻野生型小鼠相比,敲除Nrf2小鼠的过氧化氢酶活性较低,而总SOD活性未改变(图4)。
线粒体功能和动力学
为了检测线粒体含量,我们首先测量了CS和COX的最大活性,代表了TCA循环和电子传递链。老年野生型和敲除Nrf2小鼠的COX活性较低,而各组间CS活性无差异(图4)。接下来,我们测量线粒体呼吸和ROS产生作为线粒体功能的指标。随着年龄的增长,线粒体呼吸减少,而老年敲除Nrf2小鼠的ROS产生更高(图5A和B)。为了进一步评估线粒体质量,我们评估了线粒体动力学调节基因的表达。老年肌肉中Drp1 mRNA较高,而老年敲除Nrf2小鼠中Mfn1和Mfn2 mRNA较低(图6)。在蛋白质水平,敲除Nrf2导致线粒体复合物I和II减少,而III,IV和V保持不变(图7A)。虽然Mfn2蛋白含量的下降接近显着性(P = 0.10),但Nrf2缺乏对线粒体融合和裂变蛋白没有影响(图7B)。
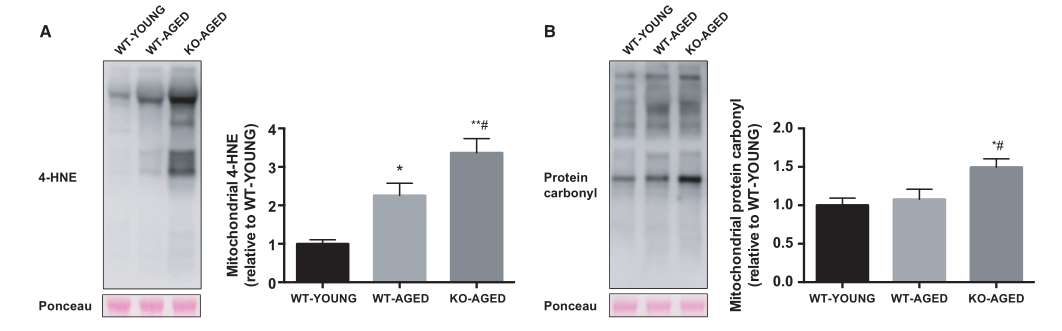
图3.老年敲除Nrf2小鼠的线粒体氧化应激。 A)线粒体4-HNE含量。 B)线粒体蛋白质羰基。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05 ** P <0.01,与年轻野生型有显著差异。 #P <0.05,与老年野生型的差异。WT,野生型;KO,淘汰赛
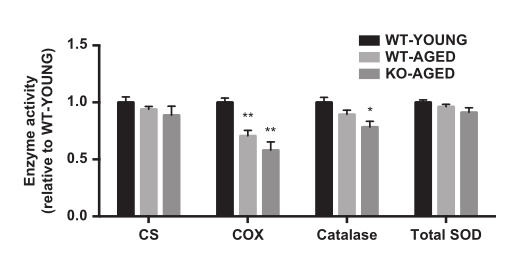
图4.老年敲除Nrf2小鼠中的酶活性。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05 ** P <0.01,与年轻野生型有显着差异。 WT,野生型; KO,淘汰赛。
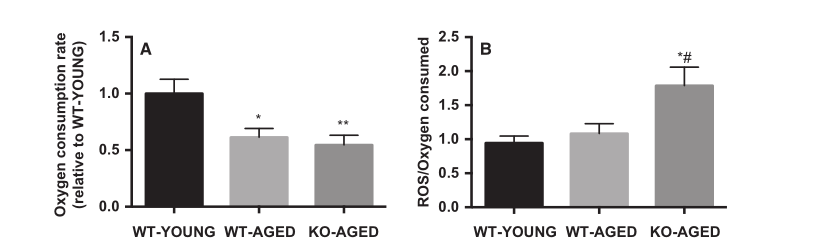
图5.老年敲除Nrf2小鼠的线粒体功能。 A)线粒体呼吸期间的耗氧率。 B)每消耗氧气的线粒体ROS产生。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05 ** P <0.01,与年轻野生型有显着差异。 #P <0.05,与老年野生型的差异。 WT,野生型; KO,淘汰赛。
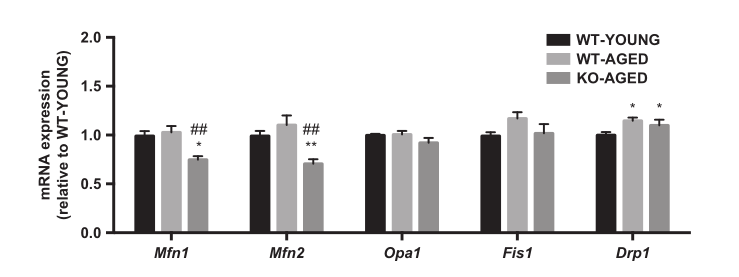
图6.老年敲除Nrf2小鼠中的线粒体动力学基因。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05,与年轻野生型有显着差异。 ## P <0.01,与老年野生型的差异。WT,野生型;KO,淘汰赛。
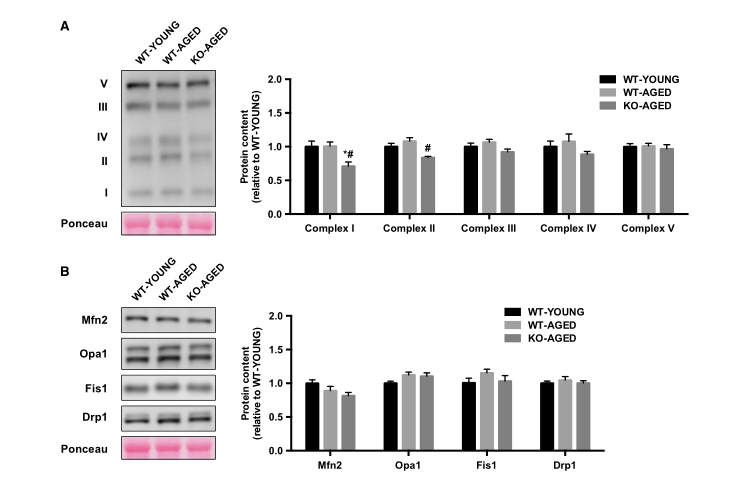
图7.来自老年敲除Nrf2小鼠的全肌裂解物中的线粒体蛋白水平。 A)线粒体OXPHOS蛋白。 B)线粒体融合和裂变调节蛋白。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05,与WTYOUNG有显着差异。 #P <0.05,与老年的差异。 WT,野生型; KO,淘汰赛。
讨论
尽管线粒体被认为是肌肉减少症的主要潜在介质,但骨骼肌线粒体与肌肉减少症之间关系的分子基础仍不清楚。衰老对线粒体含量的影响的假设一直存在争议;然而,大多数报告显示随着衰老而ROS产生增加(Carter等2015; Holloway等2018)。鉴于线粒体不仅是ROS的主要来源,而且也是氧化损伤的目标,ROS可能会产生反馈环,从而加剧线粒体功能障碍。以前的研究调查了Nrf2与线粒体功能之间的关系(Holmstrom等,2013; Coleman等,2018)。 Nrf2的缺失导致小鼠胚胎成纤维细胞(Holmstrom等2013)或UCP1转基因小鼠的肌纤维中的线粒体呼吸受损(Coleman等,2018);然而,据我们所知,尚未检查来自老年敲除Nrf2 骨骼肌的分离的线粒体组分。在这项研究中,我们发现从老化敲除Nrf2的肌肉中分离的线粒体部分中的氧化应激显著增加。如早期研究中所观察到的,Nrf2缺乏诱导其靶抗氧化基因表达的降低(Miller等,2012; Kitaoka等,2016)。重要的是,先前的研究证实了老年敲除Nrf2小鼠的整个肌肉匀浆中ROS和4-HNE水平的增加(Miller等,2012; Narasimhan等,2014)。目前使用分离的线粒体组分的研究表明氧化还原平衡被改变为氧化应激的积累,氧化应激被认为是线粒体毒性的指标。然而,尽管线粒体氧化应激升高,但Nrf2缺乏对老年骨骼肌中线粒体呼吸和肌肉减少指数没有影响。最后,我们评估了与线粒体融合和裂变相关的基因的mRNA和蛋白质表达,这是维持功能性线粒体的重要过程(Yan等,2012)。形态学上,衰老诱导骨骼肌中的线粒体断裂或异常增大(Iqbal等2013; Leduc-Gaudet等2015)。我们观察到线粒体融合调节基因在老年Nrf2肌肉中适度下调,支持ROS诱导线粒体断裂的概念(Fan等,2010; Iqbal和Hood,2014)。然而,在蛋白质水平,Mfn2与Nrf2 KO的下降没有达到显著性。需要进一步研究通过电子显微镜以检查老年敲除Nrf2肌肉中的线粒体是否显示异常超微结构。总之,我们的研究结果表明,氧化应激不是肌肉萎缩的直接原因。我们的数据与先前使用敲除Nrf2小鼠在去神经支配诱导的肌肉萎缩(Kitaoka等2016)和链脲佐菌素诱导的糖尿病萎缩模型(Whitman等2013)中的观察一致。
使用具有遗传增强的线粒体抗氧化活性的小鼠(Umanskaya等,2014)或用线粒体保护性肽治疗的小鼠(Siegel等,2013)的证据证明支持线粒体氧化损伤在年龄相关的肌肉功能障碍中的作用。此外,称为Nrf2激活剂的萝卜硫素的使用已经显示出在小鼠肌营养不良模型中改善的肌肉功能(Sun等,2015)。这项研究的局限性在于我们没有测量骨骼肌纤维的大小/数量和收缩功能。 Crilly等(2016)报道,与野生型动物相比,敲除Nrf2小鼠在分离的肌肉中表现出更高的疲劳率。最近,Ahn等(2018)报道,与年龄匹配的野生型小鼠相比,在老年敲除Nrf2小鼠中,肌肉横截面积归一化的力产生减少。这些研究表明,由于缺乏Nrf2,高水平的ROS暴露可能改变肌肉收缩功能,不一定伴随肌肉质量的变化。
在这项研究中,我们试图检查Nrf2缺乏对老年骨骼肌线粒体的影响。我们证明Nrf2缺乏增强了老年骨骼肌中线粒体ROS的产生,并加剧了与年龄相关的氧化应激,但对线粒体功能或肌肉质量几乎没有影响。
利益冲突
没有利益冲突。
参考文献(略)
翻译:
Nrf2缺乏对老年骨骼肌线粒体氧化应激的影响
Yu Kitaoka 1 , Yuki Tamura 2 , Kenya Takahashi 3 , Kohei Takeda 4 , Tohru Takemasa 4 & Hideo Hatta3
1日本横滨神奈川大学人文科学系Yu Kitaoka 1 , Yuki Tamura 2 , Kenya Takahashi 3 , Kohei Takeda 4 , Tohru Takemasa 4 & Hideo Hatta3
2日本东京日本体育科学大学运动生理学系
3日本东京大学体育科学系
4日本筑波大学综合人类科学研究科
关键词 衰老,线粒体,氧化应激,骨骼肌肉
摘要 氧化应激和线粒体功能障碍与衰老过程有关。然而,核因子红细胞系2相关因子2(Nrf2)在衰老过程中对骨骼肌的作用仍有待阐明。在目前的研究中,我们评估了缺乏Nrf2,它被称为氧化还原稳态的主要调节因子,是否会促进骨骼肌中与年龄相关的线粒体功能障碍和肌肉萎缩。在这里,我们揭示了在老年Nrf2敲除的(KO)小鼠中,由于Nrf2-靶向抗氧化基因的表达降低,线粒体中氧化应激的标志物4-羟基壬烯醛和蛋白质羰基强烈升高。线粒体呼吸随着衰老而下降,然而,敲除Nrf2 和年龄匹配的野生型小鼠之间没有差异。类似地,与年轻野生型小鼠相比,老年野生型和敲除Nrf2 小鼠中的细胞色素c氧化酶活性较低。老年敲除Nrf2 小鼠肌肉中Mfn1和Mfn2 mRNA的表达较低。在老化的敲除Nrf2小鼠中,每消耗氧气的线粒体活性氧产生增加。敲除Nrf2 对体重归一化的肌肉质量没有影响。这些结果表明,Nrf2缺乏会加剧与年龄相关的线粒体氧化应激,但不会影响骨骼肌呼吸功能的下降。
引言
衰老的特征在于骨骼肌质量和功能的丧失,这种现象称为肌肉减少症。这种肌肉功能的丧失导致身体虚弱和死亡的风险增加(Cruz-Jentoft等,2010)。因此,了解肌肉萎缩随老化的机制对于识别治疗目标和促进健康非常重要。衰老过程被认为是由氧化应激驱动的,最初由Harman 于1956年提出作为自由基理论。鉴于线粒体电子传递链是活性氧(ROS)的主要来源,线粒体被认为在肌肉减少症中起着至关重要的作用。与该假设一致,许多研究表明老年骨骼肌中ROS产生增加,与线粒体功能障碍和线粒体细胞凋亡易感性增加相关(Chabi等2008; Dai等2014)。然而,升高的ROS产生是否与肌肉萎缩和衰老有因果关系仍待澄清。
Nrf2是一种转录因子,被认为是抗氧化基因的关键调节因子(Motohashi和Yamamoto 2004)。响应于氧化应激,Nrf2易位到细胞核中并与其靶抗氧化基因的抗氧化反应元件结合。我们以前曾报道Nrf2缺乏会加重年轻小鼠骨骼肌中去神经支配诱导的氧化应激,而对肌肉质量的损失几乎没有影响(Kitaoka等,2016)。然而,据报道,Nrf2缺乏对抗氧化酶的影响在老年骨骼肌中比在幼小动物肌肉中更大(Miller等,2012)。有趣的是,Nrf2的消融导致与年龄相关的氧化应激状态中的肌肉再生受损(Narasimhan等,2014)。为了探讨Nrf2信号可能参与衰老过程,我们评估了Nrf2的缺乏是否促进骨骼肌中与年龄相关的线粒体氧化应激和肌肉萎缩。
方法
动物和实验设计
敲除Nrf2小鼠获自Jackson 实验室(Bar Harbor,ME)。如先前报道的那样通过尾部DNA的PCR分析对小鼠进行基因分型(Kitaoka等,2016)。将动物分组饲养(每笼3-4只),在空调房间中,在12小时光照,12小时黑暗环境中循环,随意给予标准饲料和水。使用老年雄性(22个月大)Nrf2敲除小鼠和年轻(4个月大)和年龄匹配(22个月大)雄性野生型C57BL / 6J(WT)小鼠(每组n = 6-7)。通过颈脱位使动物安乐死,迅速取出肌肉,快速冷冻,并在-80℃下储存。所有实验均由东京大学动物实验委员会批准。
RNA分离和实时定量PCR
在Trizol试剂(Life Technologies,Gaithersburg,MD)中将腓肠肌在冰上匀浆,然后用氯仿分离成有机相和水相。在用乙醇沉淀后,使用RNeasy Mini试剂盒(Qiagen)从水相中分离总RNA。在通过分光光度法(Nanodrop ND1000,Thermo Scientific,Waltham,MA)测量RNA浓度后,使用高容量cDNA逆转录试剂盒(Applied Biosystems,Foster City,CA)进行第一链cDNA合成。Nrf2,Nqo1(NAD(P)H醌氧化还原酶1),Cat(过氧化氢酶),Gclc(谷氨酸-半胱氨酸连接酶催化亚基),Sod1(超氧化物歧化酶1),Sod2,Fis1(裂变,线粒体1),Drp1的表达(启动相关蛋白1),Mfn1(mitofusin 1),Mfn2,Opa1(视神经萎缩1)使用Thermal Cycler Dice Real-Time System和SYBR Premix Ex taq II(Takara Bio,Shiga,Japan)定量。所有样品一式两份进行。Tbp(TATA盒结合蛋白)用作对照基因,其表达在各组之间没有改变。上述基因的正向和反向引物如表1所示。
线粒体分离
如前所述使用差速离心分离线粒体级分(Tamura等,2015)。简言之,将股四头肌肌肉在线粒体分离缓冲液(67mm蔗糖,50mm Tris,50mm KCl,10mm EDTA和0.2%(w / v)无脂肪酸的牛血清白蛋白,pH7.4)中轻度均质化。将匀浆在4℃下以700g离心15分钟,并将上清液在12,000g下离心20分钟。洗涤沉淀,并重悬于线粒体分离缓冲液中。在分离程序后,使用二辛可宁酸(BCA)蛋白质测定法(Pierce,Rockford,IL)定量样品的总蛋白质含量。线粒体样品用于分析4-HNE,蛋白质羰基,线粒体呼吸和ROS产生。我们通过蛋白质印迹使用针对甘油醛-3-磷酸脱氢酶(细胞溶质标记物)和细胞色素c氧化酶(COX)亚基IV(线粒体标记物;数据未显示)的抗体证实了线粒体部分的纯度。此外,通过在单独的实验中添加外源细胞色素来验证我们的线粒体分离方法的完整性。
整个肌肉裂解物
腓肠肌在放射免疫沉淀试验(RIPA)缓冲液(25 mmol / L Tris HCl,pH 7.6,150 mmol / L NaCl,1%NP-40,1%脱氧胆酸钠和0.1%十二烷基硫酸钠[SDS])中均质化,用蛋白酶抑制剂物(Complete Mini,ETDA-free,Roche Applied Science,Indianapolis,IN)和磷酸酶抑制剂(PhosSTOP,Roche Applied Science)混合。使用BCA蛋白质测定法(Pierce)定量样品的总蛋白质含量。
蛋白质印迹
将等量的蛋白质加载到10%SDS-PAGE凝胶上并通过电泳分离。 将蛋白质转移到聚偏二氟乙烯(PVDF)膜上,蛋白质印迹使用4-HNE(4-羟基壬烯醛; ab48506) ,Total OXPHOS Rodent WB Antibody Cocktail (ab110413),Fis1(ab96764),Drp1(ab56788),Mfn2(ab 124773) 来自美国剑桥;Opa1(#612606)来自日本东京。胭脂红染色用于验证一致负荷。使用C印迹扫描仪(LI-COR,Lincoln,NE)扫描并定量印迹。
蛋白质羰基含量
用市售试剂盒(#ROIK03; SHIMA 实验室,东京,日本)测量蛋白质羰基含量。在如上所述将线粒体蛋白转移至PVDF膜后,使膜与二硝基苯肼(DNPH)反应,然后进行Western印迹。
酶活性
将胫骨前肌在100(v / w)的100mmol / L磷酸钾缓冲液中匀浆。按照已建立的方案(Spinazzi等,2012),通过分光光度法测量柠檬酸合酶(CS)和COX的最大活性。如前所述(Hadwan,2018)使用分光光度法测定过氧化氢酶活性。使用Superoxide Dismutase Assay Kit(706002,Cayman,Ann Arbor,MI)按照制造商的说明测定总SOD(Mn-SOD和Cu / Zn-SOD)活性。
线粒体呼吸
将新鲜分离的线粒体(60μg)在反应缓冲液(250mmol / L蔗糖,10mmol / L Tris,1mmol / L MgCl 2)中温育。 使用具有MitoXpress Xtra荧光传感器试剂(Agilent Technology,Santa Clara,CA)的Tecan Spark多模板读数器测量线粒体氧消耗,以测量溶解氧水平(Ex:380nm / Em:670nm)。通过添加10mmol / L琥珀酸盐和1mol / L鱼藤酮和2.5mmol / L ADP来刺激复合物II驱动的III期呼吸。使用操作软件计算每分钟的相对荧光变化。
线粒体活性氧生成
将新鲜分离的线粒体(20μg)在线粒体呼吸缓冲液和50μl/ L 2'7'-二氯荧光素(DCF)中孵育。通过添加10mmol / L琥珀酸盐和11mol / L鱼藤酮和2.5mmol / L ADP,在状态III呼吸条件下测量ROS发射。使用Tecan多模板读数器测量相对荧光变化(Ex:480nm / Em:520nm)。
统计分析
数据表示为平均值±标准误差(SEM)。进行单向方差分析(ANOVA),然后进行Bonferroni多重比较试验(GraphPad Prism 6.0,La Jolla,CA)。统计学显着性定义为P <0.05。
结果
骨骼肌质量和肌肉减少指数
动物特征如图1所示。老年敲除Nrf2小鼠体重比老年野生型小鼠轻(图1A)。为了研究衰老和Nrf2缺乏对骨骼肌质量的影响,我们测量了腓肠肌和胫骨前肌的绝对质量,以及肌肉减少指数(肌肉质量/体重)。与老年野生型小鼠相比,老年敲除Nrf2小鼠的绝对肌肉质量较低(图1B)。衰老降低肌肉减少指数,然而,Nrf2缺乏没有影响(图1C)。
图1 老年敲除Nrf2小鼠的体重,骨骼肌质量和肌肉减少指数。 (A)体重。(B)腓肠肌和胫骨前肌肉质量。 (C)肌肉减少指数(每体重肌肉质量)。 数据表示为平均值±SEM。每组n = 6-7。* P <0.05 ** P <0.01,与年轻野生型有显着差异。#P <0.05 ## P <0.01,与老年野生型相比有显着差异。WT,野生型; KO,敲除。
图2.老年敲除Nrf2小鼠中的Nrf2靶抗氧化基因。 数据表示为平均值±每组SEM n = 5-7。* P <0.05 ** P <0.01,与年轻野生型有显着差异。#P <0.05 ## P <0.01,与老年野生型的差异。 WT,野生型;KO,敲除。
抗氧化基因表达和氧化应激
与野生型小鼠相比,通过不可检测的mRNA表达证实Nrf2的敲除。衰老对Nrf2及其主要靶基因(Nqo1,Cat,Gclc,Sod1和Sod2)的mRNA表达没有影响。Nrf2靶向抗氧化基因的表达在老年敲除Nrf2肌肉中减少,除Sod2,其他没有改变(图2)。为了检测线粒体氧化损伤,我们测量了线粒体组分中4-HNE(脂质过氧化标记物)和蛋白质羰基含量(蛋白质氧化标记物)的水平。敲除Nrf2小鼠显示4-HNE(图3A)和蛋白质羰基含量显着增加(图3B)。与年轻野生型小鼠相比,敲除Nrf2小鼠的过氧化氢酶活性较低,而总SOD活性未改变(图4)。
线粒体功能和动力学
为了检测线粒体含量,我们首先测量了CS和COX的最大活性,代表了TCA循环和电子传递链。老年野生型和敲除Nrf2小鼠的COX活性较低,而各组间CS活性无差异(图4)。接下来,我们测量线粒体呼吸和ROS产生作为线粒体功能的指标。随着年龄的增长,线粒体呼吸减少,而老年敲除Nrf2小鼠的ROS产生更高(图5A和B)。为了进一步评估线粒体质量,我们评估了线粒体动力学调节基因的表达。老年肌肉中Drp1 mRNA较高,而老年敲除Nrf2小鼠中Mfn1和Mfn2 mRNA较低(图6)。在蛋白质水平,敲除Nrf2导致线粒体复合物I和II减少,而III,IV和V保持不变(图7A)。虽然Mfn2蛋白含量的下降接近显着性(P = 0.10),但Nrf2缺乏对线粒体融合和裂变蛋白没有影响(图7B)。
图3.老年敲除Nrf2小鼠的线粒体氧化应激。 A)线粒体4-HNE含量。 B)线粒体蛋白质羰基。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05 ** P <0.01,与年轻野生型有显著差异。 #P <0.05,与老年野生型的差异。WT,野生型;KO,淘汰赛
图4.老年敲除Nrf2小鼠中的酶活性。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05 ** P <0.01,与年轻野生型有显着差异。 WT,野生型; KO,淘汰赛。
图5.老年敲除Nrf2小鼠的线粒体功能。 A)线粒体呼吸期间的耗氧率。 B)每消耗氧气的线粒体ROS产生。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05 ** P <0.01,与年轻野生型有显着差异。 #P <0.05,与老年野生型的差异。 WT,野生型; KO,淘汰赛。
图6.老年敲除Nrf2小鼠中的线粒体动力学基因。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05,与年轻野生型有显着差异。 ## P <0.01,与老年野生型的差异。WT,野生型;KO,淘汰赛。
图7.来自老年敲除Nrf2小鼠的全肌裂解物中的线粒体蛋白水平。 A)线粒体OXPHOS蛋白。 B)线粒体融合和裂变调节蛋白。 数据表示为平均值±每组SEM n = 5-7。 * P <0.05,与WTYOUNG有显着差异。 #P <0.05,与老年的差异。 WT,野生型; KO,淘汰赛。
讨论
尽管线粒体被认为是肌肉减少症的主要潜在介质,但骨骼肌线粒体与肌肉减少症之间关系的分子基础仍不清楚。衰老对线粒体含量的影响的假设一直存在争议;然而,大多数报告显示随着衰老而ROS产生增加(Carter等2015; Holloway等2018)。鉴于线粒体不仅是ROS的主要来源,而且也是氧化损伤的目标,ROS可能会产生反馈环,从而加剧线粒体功能障碍。以前的研究调查了Nrf2与线粒体功能之间的关系(Holmstrom等,2013; Coleman等,2018)。 Nrf2的缺失导致小鼠胚胎成纤维细胞(Holmstrom等2013)或UCP1转基因小鼠的肌纤维中的线粒体呼吸受损(Coleman等,2018);然而,据我们所知,尚未检查来自老年敲除Nrf2 骨骼肌的分离的线粒体组分。在这项研究中,我们发现从老化敲除Nrf2的肌肉中分离的线粒体部分中的氧化应激显著增加。如早期研究中所观察到的,Nrf2缺乏诱导其靶抗氧化基因表达的降低(Miller等,2012; Kitaoka等,2016)。重要的是,先前的研究证实了老年敲除Nrf2小鼠的整个肌肉匀浆中ROS和4-HNE水平的增加(Miller等,2012; Narasimhan等,2014)。目前使用分离的线粒体组分的研究表明氧化还原平衡被改变为氧化应激的积累,氧化应激被认为是线粒体毒性的指标。然而,尽管线粒体氧化应激升高,但Nrf2缺乏对老年骨骼肌中线粒体呼吸和肌肉减少指数没有影响。最后,我们评估了与线粒体融合和裂变相关的基因的mRNA和蛋白质表达,这是维持功能性线粒体的重要过程(Yan等,2012)。形态学上,衰老诱导骨骼肌中的线粒体断裂或异常增大(Iqbal等2013; Leduc-Gaudet等2015)。我们观察到线粒体融合调节基因在老年Nrf2肌肉中适度下调,支持ROS诱导线粒体断裂的概念(Fan等,2010; Iqbal和Hood,2014)。然而,在蛋白质水平,Mfn2与Nrf2 KO的下降没有达到显著性。需要进一步研究通过电子显微镜以检查老年敲除Nrf2肌肉中的线粒体是否显示异常超微结构。总之,我们的研究结果表明,氧化应激不是肌肉萎缩的直接原因。我们的数据与先前使用敲除Nrf2小鼠在去神经支配诱导的肌肉萎缩(Kitaoka等2016)和链脲佐菌素诱导的糖尿病萎缩模型(Whitman等2013)中的观察一致。
使用具有遗传增强的线粒体抗氧化活性的小鼠(Umanskaya等,2014)或用线粒体保护性肽治疗的小鼠(Siegel等,2013)的证据证明支持线粒体氧化损伤在年龄相关的肌肉功能障碍中的作用。此外,称为Nrf2激活剂的萝卜硫素的使用已经显示出在小鼠肌营养不良模型中改善的肌肉功能(Sun等,2015)。这项研究的局限性在于我们没有测量骨骼肌纤维的大小/数量和收缩功能。 Crilly等(2016)报道,与野生型动物相比,敲除Nrf2小鼠在分离的肌肉中表现出更高的疲劳率。最近,Ahn等(2018)报道,与年龄匹配的野生型小鼠相比,在老年敲除Nrf2小鼠中,肌肉横截面积归一化的力产生减少。这些研究表明,由于缺乏Nrf2,高水平的ROS暴露可能改变肌肉收缩功能,不一定伴随肌肉质量的变化。
在这项研究中,我们试图检查Nrf2缺乏对老年骨骼肌线粒体的影响。我们证明Nrf2缺乏增强了老年骨骼肌中线粒体ROS的产生,并加剧了与年龄相关的氧化应激,但对线粒体功能或肌肉质量几乎没有影响。
利益冲突
没有利益冲突。
参考文献(略)
本文由福山生物整理翻译,转载请注明出处。