






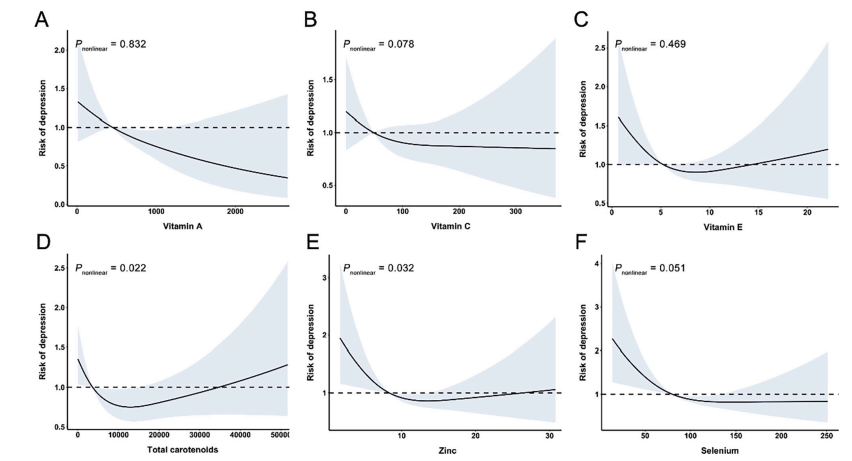
最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
核因子NF-E2相关因子(Nrf2)激活后通过调节骨骼肌氧化还原增加运动耐力
发表于:2021-05-13 作者:admin 来源:本站 点击量:19695
原文:Oh S , Komine S , Warabi E , et al. Nuclear factor (erythroid derived 2)-like 2 activation increases exercise endurance capacity via redox modulation in skeletal muscles[J]. Rep, 2017, 7(1):12902.
翻译:
萝卜硫素(SFN)通过激活核因子NF-E2相关因子(Nrf2)信号通路,在预防氧化应激中发挥重要作用。SFN可通过对抗运动过程中氧化应激引起的损伤提高运动耐力。我们通过力竭跑步测试(连续渐进式力竭测试)评估跑步能力,并检测氧化应激和肌肉损伤标志物的表达。在测试之前,给予12至13周龄雄性野生型(Nrf2 +/+)和Nrf2缺失(Nrf2 -/-)C57BL / 6J小鼠SFN腹膜注射或溶媒注射。注射SFN的Nrf2+/+小鼠奔跑距离明显大于未注射SFN的小鼠。随着Nrf2信号和下游基因的上调,运动能力增强。实验后,注射SFN的Nrf2+/+小鼠氧化应激标志物明显低于未注射SFN的小鼠。在力竭运动条件下,SFN对Nrf2+/+小鼠肌肉损伤的保护作用强于Nrf2 -/-小鼠。SFN诱导的Nrf2上调及其抗氧化作用可能通过减少过度运动引起的氧化应激在减轻肌肉疲劳方面发挥重要作用。这进而提高了运动耐力。这些结果为SFN诱导的Nrf2上调及其在改善运动表现中的作用提供了新的见解。
肌肉收缩过程中活性氧(ROS)的产生增加(1,2)。虽然生物系统能够对活性氧及其反应中间体进行解毒,但在肌肉重复或长时间收缩期间,活性氧水平的持续升高可能会导致正常氧化还原状态的失衡,从而产生氧化应激(3)。力竭运动训练尤其能促进ROS的产生(4)。长时间的运动导致骨骼肌结构损伤和收缩功能障碍(3,5)。以这种方式,骨骼肌氧化还原紊乱导致进行性肌肉无力和疲劳(即力量减小和收缩速度减慢)。这些现象表现为运动耐力下降(2,3,5-7)。
最近的研究强调了核因子NF-E2相关因子 (Nrf2)在抗氧化反应元件(ARE)驱动的内源性抗氧化/外源性解毒酶表达中的作用,以及Nrf2在减轻氧化损伤中的作用。调节NRF2相关的抗氧化信号可以保护组织和器官的氧化还原和功能稳态(9)。
萝卜硫素(SFN)是十字花科植物产生的一种化学预防化合物,是Nrf2的有效激活剂(10)。许多研究表明,SFN诱导的Nrf2激活可能对氧化应激损伤的各种器官发挥细胞保护作用(11)。Malaguti等报道SFN通过Nrf2-ARE通路调节肌肉氧化还原环境,减轻骨骼肌损伤(12)。SFN的抗氧化活性可能通过减少运动引起的氧化应激性肌肉疲劳,从而提高运动耐力(2,3,5-7)。
本研究利用SFN激活小鼠的Nrf2-ARE通路。我们评估了Nrf2激活对运动耐力的影响,方法是通过对小鼠进行力竭性(渐进性-持续性)跑步试验来估计它们的跑步距离。然后检测小鼠肌肉中氧化应激和组织损伤标志物的表达。
结果
基线评估。右下肢深部腓肠肌中央部分肌肉纤维的ATP酶染色横截面分别来自SFN注射或溶媒注射的Nrf2 +/+和Nrf2 -/-小鼠(图2A)。在Nrf2 +/+和Nrf2 -/- 小鼠中,I型和II型纤维的直径没有显著差异。四次SFN注射后纤维直径没有变化(图2B)。 Nrf2 +/+和Nrf2 -/-小鼠之间I型与II型肌肉比率的比值没有显著差异。在四次SFN注射后,这些比率没有显著改变(图2C)。各组各暗、光周期的能量消耗(图2D)和呼吸商(RQ)(图2E)无显著差异。各组ATP(图2F)、糖原(图2G)、cAMP(图2H)平均水平无显著差异。这些数据表明Nrf2基因型和SFN注射在基线时对肌肉纤维形态、生理或代谢无明显影响。
线粒体生物发生。图3显示了基线时腓肠肌匀浆中线粒体生物发生生物标志物。蛋白质印迹分析显示,在四次SFN注射后,磷酸化AMPKα在Nrf2 +/+ [SFN]小鼠中显著增加(图3A)。然而,所有组中SirT1和PGC1α的蛋白质表达没有显著差异。图3B显示线粒体生物发生标志物的mRNA表达,包括NRF-1、TFAM、p53R2、COX IV、SCO1和SCO2。在任何组中均未检测到mRNA表达升高。相对于nDNA, mtDNA拷贝数升高。这被认为是线粒体生物发生的良好标志物(15)。在Nrf2+/+小鼠和Nrf2 -/-组第四次注射SFN后,mtDNA拷贝数无显著差异(图3C)。这些数据表明,激活线粒体生物发生信号与SFN注射之间的联系很弱,而SFN注射仅进行了四次。
发光活性。图4A显示在转基因OKD48(Keap1依赖性氧化应激检测器,No-48-荧光素酶)小鼠中SFN注射期间Nrf2应答的实时体内成像。OKD48小鼠表达Nrf2-Luc融合蛋白N-末端一半,由基于Nrf2的3xAre启动子驱动(13)。在Nrf2 +/+ [CON]和Nrf2 -/- [CON]小鼠中未检测到Nrf2-Luc活化。在力竭平板试验前不久,在第四次SFN注射(75小时)后3小时,在Nrf2 +/+ [SFN]小鼠中观察到显著的Nrf2上调[图4A(a)和(b)]。在SFN注射期间,在Nrf2 +/+ [SFN]小鼠中Nrf2-Luc活化在基线、24、48、72和75小时逐渐增加[图S1(a),(b),(c),(d)和(e)]。Nrf2-Luc在Nrf2 +/+ [CON]和Nrf2 -/- [CON]组中未被激活。腓肠肌的荧光素酶活性如图4B所示。Nrf2-Luc在75小时Nrf2 +/+ [SFN]小鼠的肌肉中上调。这一变化明显高于其他组(P <0.01)。这些结果表明SFN预处理上调Nrf2。
Nrf2靶基因。图4C,D显示所选Nrf2靶基因的mRNA水平:血红素加氧酶-1(HO-1)、NAD(P)H:醌氧化还原酶A(NQO1)、γ-谷氨酰半胱氨酸合成酶(γ-GCS)和过氧化氢酶。在有和没有SFN预处理的腓肠肌(图4C)和比目鱼肌(图4D)组织匀浆中检测这些基因mRNA水平。与Nrf2 -/- 组(Nrf2 -/- [SFN]和Nrf2 -/- [CON])相比,Nrf2 +/+ [SFN]小鼠在腓肠肌和比目鱼肌中所有靶基因的表达水平显著增高。在两个Nrf2 +/+组之间,腓肠肌过氧化氢酶没有上调。与Nrf2+/+[CON]小鼠相比,Nrf2+/+[SFN]小鼠比目鱼中HO-1没有上调。然而,Nrf2 +/+ [SFN]小鼠组织中其他Nrf2靶基因的表达水平显著高于Nrf2 +/+ [CON]:Nrf2 +/+[CON]小鼠腓肠肌和比目鱼肌的NQO1和γ-GCS表达水平显著高于Nrf2 -/-小鼠。
运动耐力和能量代谢参数。图5A为跑步距离测试结果,该测试作为运动耐力的评价指标。Nrf2+/+小鼠比Nrf2 -/-小鼠跑得更远。在Nrf2+/+组中,Nrf2 +/+[SFN]小鼠的跑步距离大于Nrf2 +/+[CON]小鼠。图5B为每组跑步期间耗氧量(VO2)和RQ,以及记录的跑步距离中值。图5C和D显示了在50分钟的跑步机测试中,各组的平均VO2和RQ水平。运动40分钟后各组VO2水平无显著差异。在41 - 50min期间,Nrf2 -/-组VO2水平高于Nrf2 +/+组。但四组间的平均RQ在时间周期上无显著差异。在基线和50min后,四组的血清葡萄糖和游离脂肪酸(FFAs)水平无显著差异(图5E和F)。
氧化应激和肌肉损伤的生物标志物。图6A显示在测试结束后50分钟和18小时的彻底测试后,基线时腓肠肌匀浆中硫代巴比妥酸反应物质(TBARS)的水平。在基线和50分钟期间,所有组显示出相当的TBARS水平。然而,在试验后18小时,Nrf2 +/+小鼠的TBARS水平明显低于Nrf2 -/-小鼠。SFN注射Nrf2 +/+小鼠的TBARS水平低于未注射Nrf2 +/+小鼠。对于基线的GSSG/GSH比值,所有组也显示出可比性。然而,在试验结束后50分钟,Nrf2 +/+小鼠注射SFN后,GSSG/GSH比值低于Nrf2 -/-组(图6A)。图6B显示了在基线和跑步机测试50分钟后各组肌肉损伤标志物肌酸磷酸激酶(CPK)和乳酸脱氢酶(LDH)的血清水平。在基线时,Nrf2 +/+组的CPK水平明显高于Nrf2 -/-组。在跑步机疲劳试验50分钟时,Nrf2 +/+组的CPK水平低于Nrf2 -/-组。特别是Nrf2 +/+[SFN]小鼠的CPK水平明显低于其他干预组。所有组在基线时的LDH水平相当。然而,在50分钟跑步机试验后,Nrf2 +/+组的LDH水平低于Nrf2 -/-组。在基线时,Nrf2 -/-组的血乳酸水平明显低于Nrf2 +/+组。在注射SFN 50分钟后,我们没有发现Nrf2+/+小鼠血乳酸水平显著升高。然而,其他组有显著的增加(图6C)。
这些数据表明,在力竭运动条件下,Nrf2 +/+小鼠注射SFN对肌肉有保护作用。
讨论
随着运动强度的增加,肌肉对氧的利用率提高,可能会促进线粒体电子传递链中电子溢出反应产生自由基和其他活性氧(5)。活跃肌肉中自由基形成的增强可调节多种细胞信号通路,从而可能导致肌肉收缩受限(16)。这些过程可能是由中枢神经系统功能受损、肌膜功能紊乱、微血管调节、钙调节、肌丝收缩受损和/或线粒体代谢改变引起的(17)。尽管存在争议(18,19),越来越多的证据表明,通过补充营养分子来增强身体的抗氧化防御系统,并作为活性氧清除剂,可以预防运动引起的氧化应激,减少肌肉损伤(20-22)。考虑到对Nrf2的激活,SFN是一种非常有效的抗氧化剂(11)。
我们的研究表明,SFN预处理可以通过抑制骨骼肌中TBARS的产生、GSSG/GSH的比值 (图6A),以及抑制血液中CPK、LDH(图6B)和乳酸(图6C)的释放,从而增加跑步机疲劳试验中的跑步距离(图5A)。TBARS水平升高表明体内脂质过氧化作用增加。血清CPK和LDH水平常被用来测量肌肉细胞的结构损伤。乳酸盐的积累会导致肌肉疲劳。据我们所知,我们的研究提供了第一个直接证据,证明SFN对Nrf2信号通路的上调以及对下游II相和抗氧化基因(HO-1、NQO1、γ-GCS和过氧化氢酶)的调节,在抵抗力竭运动介导的氧化应激和骨骼肌组织损伤方面起着至关重要的作用。SFN诱导的骨骼肌Nrf2激活可能在提高运动耐力方面发挥关键作用。
本研究的另一个主要目标是通过Nrf2-荧光素全身成像和肌肉荧光素酶活性测定评估体内Nrf2的激活。迄今为止,利用SDS-PAGE对肌肉Nrf2进行检测的尝试已多次失败。我们通过电刺激C2C12细胞鉴定了Nrf2的体外活性(95-110 kDa带)(23),但对小鼠肌肉Nrf2活性的检测是比较困难的。一些研究将55-65kDa区域的非特异性条带定为Nrf2 (24,25)。2012年,我们首次开发了转基因小鼠(OKD48),通过测量发光活性的成像技术研究Keap1-Nrf2通路(14)。为了提高对Nrf2信号的检测,我们开发了一种新的小鼠模型,将OKD48与Nrf2 +/+ (albino-BL/6)或Nrf2 -/- (albino-Nrf2 -/-)小鼠杂交。利用这个新模型,我们证明了SFN在全身(图4A)和肌肉组织(图4B)中激活Nrf2信号通路,并对氧化应激具有很强的保护作用。
TBARS是公认的氧化应激生物学指标(26)。脂质过氧化程度反映了与氧化应激相关的主要病理和毒理学状态(27)。越来越多的证据表明,由于短时间的高强度运动,血浆和组织中的TBARS浓度都有所增加(28 - 30)。谷胱甘肽是一种抗氧化剂,被用作氧化应激的标志。在氧化应激条件下,GSH浓度较低,GSSG浓度较高,GSSG/GSH比值增加(31)。许多研究表明,GSSG/GSH比值随运动训练的增加而增加,且与乳酸/丙酮酸比值高度相关(31,32)。高水平的氧化剂可导致收缩功能障碍、线粒体功能障碍和肌肉萎缩,所有这些都会导致肌肉无力和疲劳(3,16,33)。我们的数据表明,跑步机试验后TBARS和GSSG/GSH比值浓度的降低(12),通过SFN激活Nrf2上调下游的抗氧化剂和解毒基因,可以保护肌肉免受损伤。我们对SFN预处理与氧化剂抑制之间关系的观察与最近的报道一致。Malaguti等人报道(12),在跑步机疲劳试验后,体内氧化应激水平增加,但SFN预处理后显著降低。Angeloni等人证明SFN显著增加体外抗氧化活性(34)。
Nrf2除了减轻氧化应激外,还可能在脂质和葡萄糖代谢中发挥重要作用(35 - 37)。张和同事们的研究表明,SFN增强Nrf2表达、激活肝脏激酶B1/AMPK途径和下游基因,如过氧化物酶体增殖物激活受体γ共激活因子-1、磷酸化乙酰辅酶A羧化酶、肉碱棕榈酰转移酶-1(38)。已知AMPK在肌肉中受到调节,并且通过促进脂肪酸摄取和氧化、葡萄糖摄取和线粒体生物发生起作用(39,40)。此外,Uruno等人证明Nrf2在最大增量跑步机方案的情况下调节骨骼肌糖原代谢,特别是糖原分支酶(GBE1)(41)。这可能是因为SFN的增加促进了能量代谢,并增加了小鼠的跑步距离。这是一个需要进一步研究的重要课题,需要进一步的实验来加深我们对SFN影响运动耐力机制的理解。在这项研究中,VO2和RQ数据是通过评估能量底物、碳水化合物和脂肪的利用获得的。Nrf2 +/+组和Nrf2 -/-组VO2水平在41 - 50分钟内发生了显著变化(图5C)。同时Nrf2 -/-组的LDH和CPK水平升高(图6B);然而,我们没有检测到碳水化合物和脂肪氧化以及RQ的显著变化(图5D)。我们检测了血清中葡萄糖和FFAs的水平,这两种物质是骨骼肌的主要燃料来源,但我们没有发现四组之间的差异。因此,Nrf2 -/-组观察到的严重肌肉损伤可能是由于试验期间缺乏Nrf2表达,而不是由于缺乏能量补充。在Nrf2 +/+小鼠中,SFN介导的强诱导细胞和线粒体总抗氧化剂的产生,以及II相酶的产生,可能解释运动耐力的增强。骨骼肌氧化应激水平的变化可能是跑步距离增加的原因。
Sun等报道,SFN干预(2mg/kg) 8周,通过对肌营养不良(mdx)小鼠的氧化应激产生保护作用,改善了肌肉功能,减少了病理(43)。这些结果表明,注射SFN的Nrf2 +/+小鼠和Nrf2 -/-组中Nrf2的缺失可以影响肌肉成分,这反过来可能有助于观察运动耐力能力的差异。然而,在我们的研究中,注射SFN超过4天并没有改变肌肉纤维的类型和大小(图2A-C)。Nrf2基因的存在与否并不影响纤维的组成和形态。因此,我们认为在目前的力竭测试中,SFN诱导的运动能力提高并不受肌肉成分变化的调节。
我们展示了线粒体生物发生的标记物(图3),以阐明运动能力增加的原因。我们发现四次SFN注射后显著激活AMPKα (图3),与最近的研究一致(32)。然而,我们没有检测其他线粒体生物发生标志物的显著差异,包括四组中mtDNA拷贝数(图3A-C)。此外,我们评估了cAMP的变化(图2H),这被认为是线粒体生物发生的效应(44)。然而,我们没有测量各组之间cAMP的显著差异。Kitaoka等人证明Nrf2基因的存在并不影响线粒体形态的改变(45)。因此,我们假设激活线粒体生物发生信号与通过注射SFN提高运动耐力能力之间的联系较弱,而SFN注射仅进行了四次。
Nrf2 +/+和Nrf2 -/-小鼠的运动能力与一些研究报道的结果不一致(41,46)。我们认识到这种不一致是由于不同的运动耐力测试方案造成的。由于本研究最重要的目的是建立Nrf2 +/+小鼠注射SFN时运动能力和Nrf2 -/-组Nrf2缺失的差异,因此我们更重视运动耐力测试方案。通过试验性实验,我们发现运动试验方案的运动耐力能力差异很大,其中我们选择了最佳试验方案来证明我们的假设。前面提到的研究通过在跑步机上逐渐增加速度和/或倾斜至最大运动水平来测量运动能力。本研究有可能在运动耐力测试中使用厌氧动力而非有氧动力。若以厌氧动力为基础,则小鼠腿的动力无法跟随较高的速度负荷和坡度阻力。因此,遵循测试方案的研究之间结果的不一致是可以想象的。然而,我们认识到保持28米/分钟到力竭是非常适合评估运动耐力能力的。
最近,有关于ROS和肌肉运动适应的重要报道(18,19)。也就是说,抗氧化补充剂可能会妨碍运动训练引起的骨骼肌适应。因此,我们应该谨慎地得出有关长期SFN干预的结论。然而,即使抗氧化剂具有益处,但根据类型、持续时间、频率和剂量,它可能具有毒副作用。因此,我们认为需要更多科学证据证明抗氧化功能。我们不认为SFN仅仅是一种抗氧化剂。当然,很明显,通过SFN干预Nrf2活化的主要作用是间接表达具有抗氧化特性的各种转录因子。然而,难以说明萝卜硫素直接影响其他抗氧化剂以诱导肌肉适应的细胞信号传导。因此,需要更多的实验来回答这个问题,以确定SFN的长期负面影响。
我们的研究结果表明,SFN诱导的Nrf2上调和SFN介导的抗氧化作用可能通过减少力竭运动引起的氧化应激来减轻肌肉疲劳。最终可能会提高运动耐力。 我们相信这项研究为Nrf2在改善运动表现中的作用提供了新的见解。
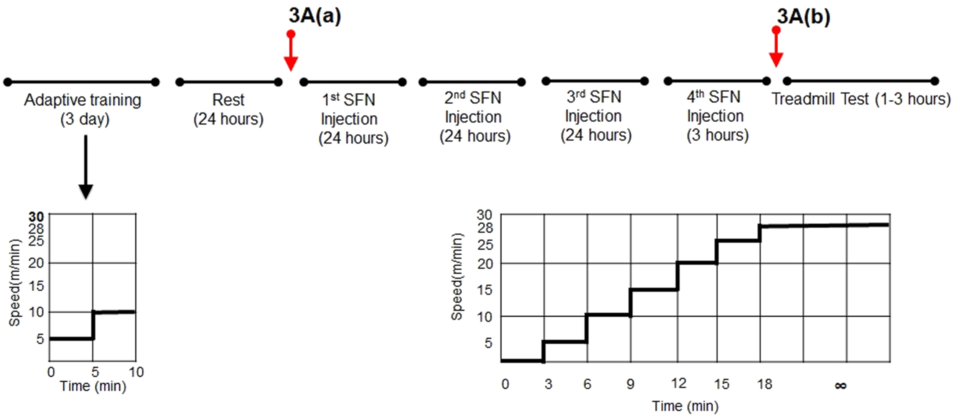
图1. 实验方案。连续3天,所有小鼠均以5-10米/分的速度低速进行10分的适应性训练。然后,所有小鼠在3天内 SFN或溶媒注射4次(72、48、24和3小时后再进行跑步机测试)。实验当天,所有小鼠均进行该实验方案。运动强度逐渐增加,初始速度为5米/分,然后每3分钟逐渐加速,至最大速度28米/分。保持最大速度,直到小鼠精疲力尽。红色箭头表示SFN或溶媒注射后体内成像的时间进程。结果如图3A[(a)~(b)]所示。

图2. SFN或溶媒注射后在pH 10.8下通过ATP酶染色获得的腓肠肌纤维类型(A)。用ATP酶染色测定腓肠肌中的纤维面积(B)和纤维类型分布(C)。使用间接量热计在每个黑暗和光照循环期间评估每日EE(D)和RQ(E)值。测量腓肠肌ATP(F)、糖原(G)和cAMP(H)水平。
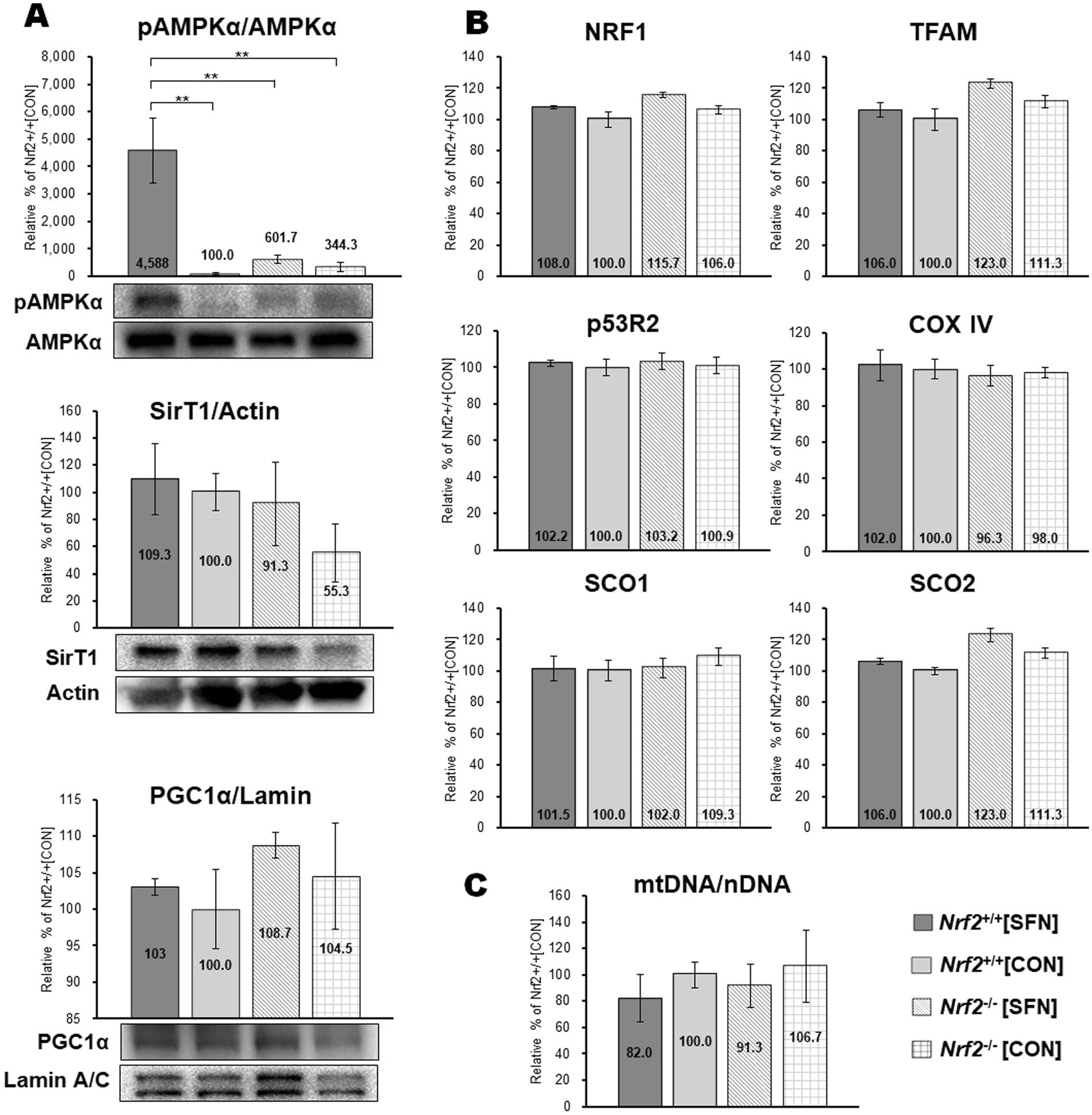
图3. SFN注射效果。SFN或溶媒注射4次,持续3天,并检测小鼠腓肠肌线粒体生物发生标志物。(A)线粒体生物发生标志物的蛋白质表达水平(蛋白质印迹条带)。 将一定量的磷酸化AMPKα标准化为AMPKα蛋白的量。还检测量了线粒体生物发生标志物SirT1和PGC1α的蛋白质含量。载荷体积分别通过肌动蛋白和核纤层蛋白A / C的表达水平标准化。(B)AA线粒体生物发生标志物的mRNA表达。通过实时PCR测定NRF1、TFAM、p53R2、COX IV、SCO1和SCO2的mRNA水平。将值标准化为管家基因GAPDH的水平。(C)腓肠肌mtDNA拷贝数。这个数字影响细胞核和mtDNA比值(mtDNA / nDNA); 括号** P <0.01。
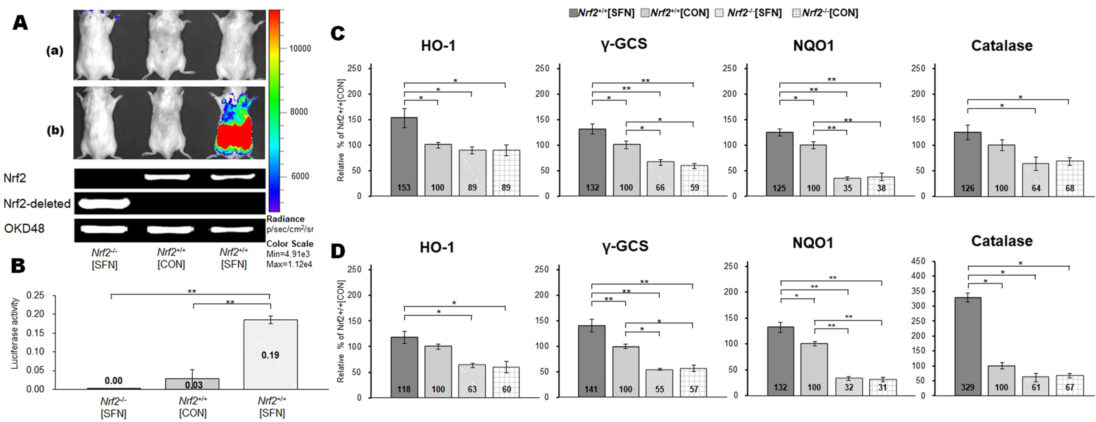
图4. 采用体内成像系统检测SFN诱导的Nrf2- luc活性,并对处于俯卧位的Nrf2和OKD48转基因小鼠(A)进行基因分型。荧光素酶测定的时间过程[(a:基线)?(b:SFN注射后)]如图1所示。使用分光计(B)测量第四次SFN注射后腓肠肌Nrf2-Luc活性。四次SFN注射或溶媒注射后,腓肠肌(C)和比目鱼肌(D)Nrf2靶基因的mRNA水平。括号* P <0.01。
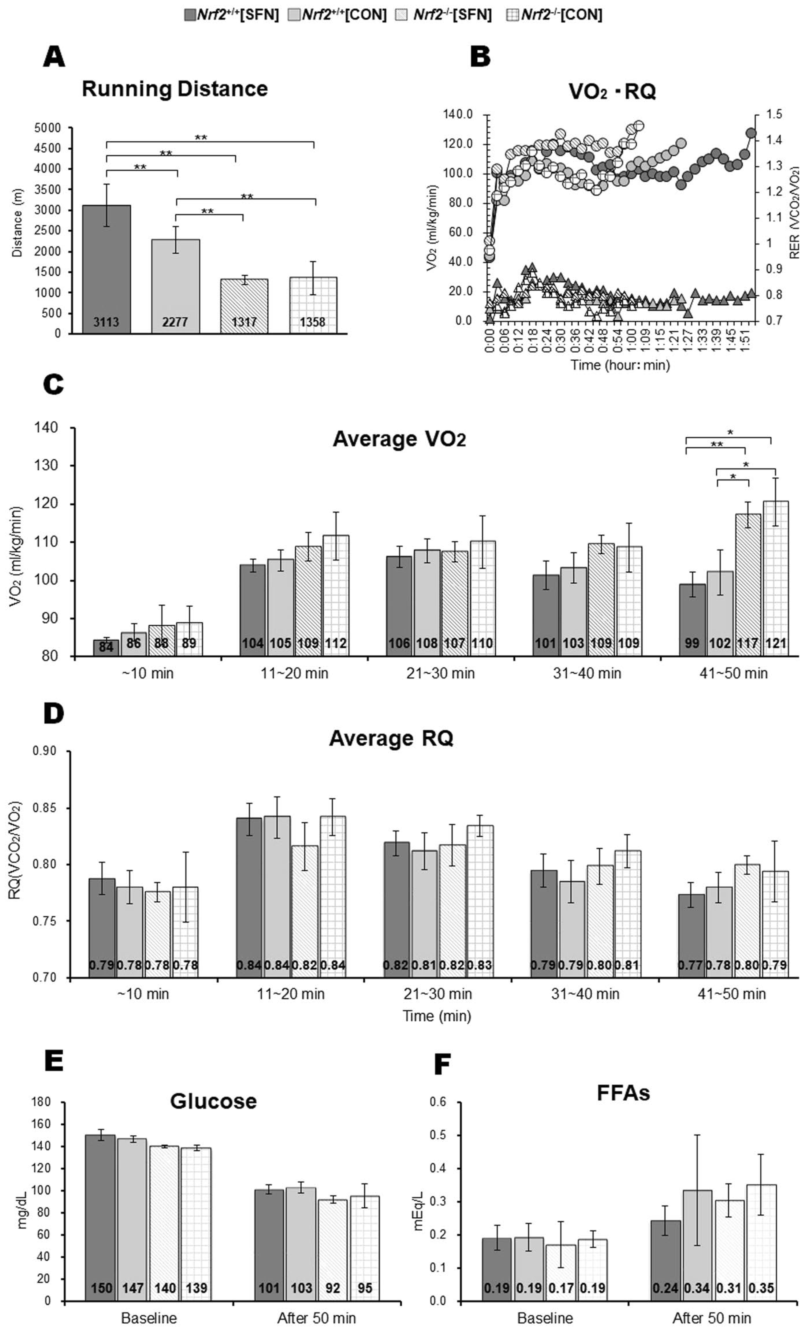
图5. 在力竭跑步试验(A)中,各组小鼠每次跑步的距离以及每次跑步期间的VO2(圆圈)和RQ(三角形)记录了跑步距离中值(B)。各组平均能量底物水平:VO2 (C)、RQ (D)、葡萄糖(E)、FFA (F) ,经过50分钟的跑步机疲劳试验后,各组在同一时间进行比较。括号* P < 0.01。

图6. 腓肠肌氧化、TBARS和GSSG/GSH (A)标记物,以及肌肉损伤、CPK和LDH (B)标记物、肌肉疲劳、血液乳酸(C)标记物。
翻译:
核因子NF-E2相关因子(Nrf2)激活后通过调节骨骼肌氧化还原增加运动耐力
摘要:萝卜硫素(SFN)通过激活核因子NF-E2相关因子(Nrf2)信号通路,在预防氧化应激中发挥重要作用。SFN可通过对抗运动过程中氧化应激引起的损伤提高运动耐力。我们通过力竭跑步测试(连续渐进式力竭测试)评估跑步能力,并检测氧化应激和肌肉损伤标志物的表达。在测试之前,给予12至13周龄雄性野生型(Nrf2 +/+)和Nrf2缺失(Nrf2 -/-)C57BL / 6J小鼠SFN腹膜注射或溶媒注射。注射SFN的Nrf2+/+小鼠奔跑距离明显大于未注射SFN的小鼠。随着Nrf2信号和下游基因的上调,运动能力增强。实验后,注射SFN的Nrf2+/+小鼠氧化应激标志物明显低于未注射SFN的小鼠。在力竭运动条件下,SFN对Nrf2+/+小鼠肌肉损伤的保护作用强于Nrf2 -/-小鼠。SFN诱导的Nrf2上调及其抗氧化作用可能通过减少过度运动引起的氧化应激在减轻肌肉疲劳方面发挥重要作用。这进而提高了运动耐力。这些结果为SFN诱导的Nrf2上调及其在改善运动表现中的作用提供了新的见解。
肌肉收缩过程中活性氧(ROS)的产生增加(1,2)。虽然生物系统能够对活性氧及其反应中间体进行解毒,但在肌肉重复或长时间收缩期间,活性氧水平的持续升高可能会导致正常氧化还原状态的失衡,从而产生氧化应激(3)。力竭运动训练尤其能促进ROS的产生(4)。长时间的运动导致骨骼肌结构损伤和收缩功能障碍(3,5)。以这种方式,骨骼肌氧化还原紊乱导致进行性肌肉无力和疲劳(即力量减小和收缩速度减慢)。这些现象表现为运动耐力下降(2,3,5-7)。
最近的研究强调了核因子NF-E2相关因子 (Nrf2)在抗氧化反应元件(ARE)驱动的内源性抗氧化/外源性解毒酶表达中的作用,以及Nrf2在减轻氧化损伤中的作用。调节NRF2相关的抗氧化信号可以保护组织和器官的氧化还原和功能稳态(9)。
萝卜硫素(SFN)是十字花科植物产生的一种化学预防化合物,是Nrf2的有效激活剂(10)。许多研究表明,SFN诱导的Nrf2激活可能对氧化应激损伤的各种器官发挥细胞保护作用(11)。Malaguti等报道SFN通过Nrf2-ARE通路调节肌肉氧化还原环境,减轻骨骼肌损伤(12)。SFN的抗氧化活性可能通过减少运动引起的氧化应激性肌肉疲劳,从而提高运动耐力(2,3,5-7)。
本研究利用SFN激活小鼠的Nrf2-ARE通路。我们评估了Nrf2激活对运动耐力的影响,方法是通过对小鼠进行力竭性(渐进性-持续性)跑步试验来估计它们的跑步距离。然后检测小鼠肌肉中氧化应激和组织损伤标志物的表达。
结果
基线评估。右下肢深部腓肠肌中央部分肌肉纤维的ATP酶染色横截面分别来自SFN注射或溶媒注射的Nrf2 +/+和Nrf2 -/-小鼠(图2A)。在Nrf2 +/+和Nrf2 -/- 小鼠中,I型和II型纤维的直径没有显著差异。四次SFN注射后纤维直径没有变化(图2B)。 Nrf2 +/+和Nrf2 -/-小鼠之间I型与II型肌肉比率的比值没有显著差异。在四次SFN注射后,这些比率没有显著改变(图2C)。各组各暗、光周期的能量消耗(图2D)和呼吸商(RQ)(图2E)无显著差异。各组ATP(图2F)、糖原(图2G)、cAMP(图2H)平均水平无显著差异。这些数据表明Nrf2基因型和SFN注射在基线时对肌肉纤维形态、生理或代谢无明显影响。
线粒体生物发生。图3显示了基线时腓肠肌匀浆中线粒体生物发生生物标志物。蛋白质印迹分析显示,在四次SFN注射后,磷酸化AMPKα在Nrf2 +/+ [SFN]小鼠中显著增加(图3A)。然而,所有组中SirT1和PGC1α的蛋白质表达没有显著差异。图3B显示线粒体生物发生标志物的mRNA表达,包括NRF-1、TFAM、p53R2、COX IV、SCO1和SCO2。在任何组中均未检测到mRNA表达升高。相对于nDNA, mtDNA拷贝数升高。这被认为是线粒体生物发生的良好标志物(15)。在Nrf2+/+小鼠和Nrf2 -/-组第四次注射SFN后,mtDNA拷贝数无显著差异(图3C)。这些数据表明,激活线粒体生物发生信号与SFN注射之间的联系很弱,而SFN注射仅进行了四次。
发光活性。图4A显示在转基因OKD48(Keap1依赖性氧化应激检测器,No-48-荧光素酶)小鼠中SFN注射期间Nrf2应答的实时体内成像。OKD48小鼠表达Nrf2-Luc融合蛋白N-末端一半,由基于Nrf2的3xAre启动子驱动(13)。在Nrf2 +/+ [CON]和Nrf2 -/- [CON]小鼠中未检测到Nrf2-Luc活化。在力竭平板试验前不久,在第四次SFN注射(75小时)后3小时,在Nrf2 +/+ [SFN]小鼠中观察到显著的Nrf2上调[图4A(a)和(b)]。在SFN注射期间,在Nrf2 +/+ [SFN]小鼠中Nrf2-Luc活化在基线、24、48、72和75小时逐渐增加[图S1(a),(b),(c),(d)和(e)]。Nrf2-Luc在Nrf2 +/+ [CON]和Nrf2 -/- [CON]组中未被激活。腓肠肌的荧光素酶活性如图4B所示。Nrf2-Luc在75小时Nrf2 +/+ [SFN]小鼠的肌肉中上调。这一变化明显高于其他组(P <0.01)。这些结果表明SFN预处理上调Nrf2。
Nrf2靶基因。图4C,D显示所选Nrf2靶基因的mRNA水平:血红素加氧酶-1(HO-1)、NAD(P)H:醌氧化还原酶A(NQO1)、γ-谷氨酰半胱氨酸合成酶(γ-GCS)和过氧化氢酶。在有和没有SFN预处理的腓肠肌(图4C)和比目鱼肌(图4D)组织匀浆中检测这些基因mRNA水平。与Nrf2 -/- 组(Nrf2 -/- [SFN]和Nrf2 -/- [CON])相比,Nrf2 +/+ [SFN]小鼠在腓肠肌和比目鱼肌中所有靶基因的表达水平显著增高。在两个Nrf2 +/+组之间,腓肠肌过氧化氢酶没有上调。与Nrf2+/+[CON]小鼠相比,Nrf2+/+[SFN]小鼠比目鱼中HO-1没有上调。然而,Nrf2 +/+ [SFN]小鼠组织中其他Nrf2靶基因的表达水平显著高于Nrf2 +/+ [CON]:Nrf2 +/+[CON]小鼠腓肠肌和比目鱼肌的NQO1和γ-GCS表达水平显著高于Nrf2 -/-小鼠。
运动耐力和能量代谢参数。图5A为跑步距离测试结果,该测试作为运动耐力的评价指标。Nrf2+/+小鼠比Nrf2 -/-小鼠跑得更远。在Nrf2+/+组中,Nrf2 +/+[SFN]小鼠的跑步距离大于Nrf2 +/+[CON]小鼠。图5B为每组跑步期间耗氧量(VO2)和RQ,以及记录的跑步距离中值。图5C和D显示了在50分钟的跑步机测试中,各组的平均VO2和RQ水平。运动40分钟后各组VO2水平无显著差异。在41 - 50min期间,Nrf2 -/-组VO2水平高于Nrf2 +/+组。但四组间的平均RQ在时间周期上无显著差异。在基线和50min后,四组的血清葡萄糖和游离脂肪酸(FFAs)水平无显著差异(图5E和F)。
氧化应激和肌肉损伤的生物标志物。图6A显示在测试结束后50分钟和18小时的彻底测试后,基线时腓肠肌匀浆中硫代巴比妥酸反应物质(TBARS)的水平。在基线和50分钟期间,所有组显示出相当的TBARS水平。然而,在试验后18小时,Nrf2 +/+小鼠的TBARS水平明显低于Nrf2 -/-小鼠。SFN注射Nrf2 +/+小鼠的TBARS水平低于未注射Nrf2 +/+小鼠。对于基线的GSSG/GSH比值,所有组也显示出可比性。然而,在试验结束后50分钟,Nrf2 +/+小鼠注射SFN后,GSSG/GSH比值低于Nrf2 -/-组(图6A)。图6B显示了在基线和跑步机测试50分钟后各组肌肉损伤标志物肌酸磷酸激酶(CPK)和乳酸脱氢酶(LDH)的血清水平。在基线时,Nrf2 +/+组的CPK水平明显高于Nrf2 -/-组。在跑步机疲劳试验50分钟时,Nrf2 +/+组的CPK水平低于Nrf2 -/-组。特别是Nrf2 +/+[SFN]小鼠的CPK水平明显低于其他干预组。所有组在基线时的LDH水平相当。然而,在50分钟跑步机试验后,Nrf2 +/+组的LDH水平低于Nrf2 -/-组。在基线时,Nrf2 -/-组的血乳酸水平明显低于Nrf2 +/+组。在注射SFN 50分钟后,我们没有发现Nrf2+/+小鼠血乳酸水平显著升高。然而,其他组有显著的增加(图6C)。
这些数据表明,在力竭运动条件下,Nrf2 +/+小鼠注射SFN对肌肉有保护作用。
讨论
随着运动强度的增加,肌肉对氧的利用率提高,可能会促进线粒体电子传递链中电子溢出反应产生自由基和其他活性氧(5)。活跃肌肉中自由基形成的增强可调节多种细胞信号通路,从而可能导致肌肉收缩受限(16)。这些过程可能是由中枢神经系统功能受损、肌膜功能紊乱、微血管调节、钙调节、肌丝收缩受损和/或线粒体代谢改变引起的(17)。尽管存在争议(18,19),越来越多的证据表明,通过补充营养分子来增强身体的抗氧化防御系统,并作为活性氧清除剂,可以预防运动引起的氧化应激,减少肌肉损伤(20-22)。考虑到对Nrf2的激活,SFN是一种非常有效的抗氧化剂(11)。
我们的研究表明,SFN预处理可以通过抑制骨骼肌中TBARS的产生、GSSG/GSH的比值 (图6A),以及抑制血液中CPK、LDH(图6B)和乳酸(图6C)的释放,从而增加跑步机疲劳试验中的跑步距离(图5A)。TBARS水平升高表明体内脂质过氧化作用增加。血清CPK和LDH水平常被用来测量肌肉细胞的结构损伤。乳酸盐的积累会导致肌肉疲劳。据我们所知,我们的研究提供了第一个直接证据,证明SFN对Nrf2信号通路的上调以及对下游II相和抗氧化基因(HO-1、NQO1、γ-GCS和过氧化氢酶)的调节,在抵抗力竭运动介导的氧化应激和骨骼肌组织损伤方面起着至关重要的作用。SFN诱导的骨骼肌Nrf2激活可能在提高运动耐力方面发挥关键作用。
本研究的另一个主要目标是通过Nrf2-荧光素全身成像和肌肉荧光素酶活性测定评估体内Nrf2的激活。迄今为止,利用SDS-PAGE对肌肉Nrf2进行检测的尝试已多次失败。我们通过电刺激C2C12细胞鉴定了Nrf2的体外活性(95-110 kDa带)(23),但对小鼠肌肉Nrf2活性的检测是比较困难的。一些研究将55-65kDa区域的非特异性条带定为Nrf2 (24,25)。2012年,我们首次开发了转基因小鼠(OKD48),通过测量发光活性的成像技术研究Keap1-Nrf2通路(14)。为了提高对Nrf2信号的检测,我们开发了一种新的小鼠模型,将OKD48与Nrf2 +/+ (albino-BL/6)或Nrf2 -/- (albino-Nrf2 -/-)小鼠杂交。利用这个新模型,我们证明了SFN在全身(图4A)和肌肉组织(图4B)中激活Nrf2信号通路,并对氧化应激具有很强的保护作用。
TBARS是公认的氧化应激生物学指标(26)。脂质过氧化程度反映了与氧化应激相关的主要病理和毒理学状态(27)。越来越多的证据表明,由于短时间的高强度运动,血浆和组织中的TBARS浓度都有所增加(28 - 30)。谷胱甘肽是一种抗氧化剂,被用作氧化应激的标志。在氧化应激条件下,GSH浓度较低,GSSG浓度较高,GSSG/GSH比值增加(31)。许多研究表明,GSSG/GSH比值随运动训练的增加而增加,且与乳酸/丙酮酸比值高度相关(31,32)。高水平的氧化剂可导致收缩功能障碍、线粒体功能障碍和肌肉萎缩,所有这些都会导致肌肉无力和疲劳(3,16,33)。我们的数据表明,跑步机试验后TBARS和GSSG/GSH比值浓度的降低(12),通过SFN激活Nrf2上调下游的抗氧化剂和解毒基因,可以保护肌肉免受损伤。我们对SFN预处理与氧化剂抑制之间关系的观察与最近的报道一致。Malaguti等人报道(12),在跑步机疲劳试验后,体内氧化应激水平增加,但SFN预处理后显著降低。Angeloni等人证明SFN显著增加体外抗氧化活性(34)。
Nrf2除了减轻氧化应激外,还可能在脂质和葡萄糖代谢中发挥重要作用(35 - 37)。张和同事们的研究表明,SFN增强Nrf2表达、激活肝脏激酶B1/AMPK途径和下游基因,如过氧化物酶体增殖物激活受体γ共激活因子-1、磷酸化乙酰辅酶A羧化酶、肉碱棕榈酰转移酶-1(38)。已知AMPK在肌肉中受到调节,并且通过促进脂肪酸摄取和氧化、葡萄糖摄取和线粒体生物发生起作用(39,40)。此外,Uruno等人证明Nrf2在最大增量跑步机方案的情况下调节骨骼肌糖原代谢,特别是糖原分支酶(GBE1)(41)。这可能是因为SFN的增加促进了能量代谢,并增加了小鼠的跑步距离。这是一个需要进一步研究的重要课题,需要进一步的实验来加深我们对SFN影响运动耐力机制的理解。在这项研究中,VO2和RQ数据是通过评估能量底物、碳水化合物和脂肪的利用获得的。Nrf2 +/+组和Nrf2 -/-组VO2水平在41 - 50分钟内发生了显著变化(图5C)。同时Nrf2 -/-组的LDH和CPK水平升高(图6B);然而,我们没有检测到碳水化合物和脂肪氧化以及RQ的显著变化(图5D)。我们检测了血清中葡萄糖和FFAs的水平,这两种物质是骨骼肌的主要燃料来源,但我们没有发现四组之间的差异。因此,Nrf2 -/-组观察到的严重肌肉损伤可能是由于试验期间缺乏Nrf2表达,而不是由于缺乏能量补充。在Nrf2 +/+小鼠中,SFN介导的强诱导细胞和线粒体总抗氧化剂的产生,以及II相酶的产生,可能解释运动耐力的增强。骨骼肌氧化应激水平的变化可能是跑步距离增加的原因。
Sun等报道,SFN干预(2mg/kg) 8周,通过对肌营养不良(mdx)小鼠的氧化应激产生保护作用,改善了肌肉功能,减少了病理(43)。这些结果表明,注射SFN的Nrf2 +/+小鼠和Nrf2 -/-组中Nrf2的缺失可以影响肌肉成分,这反过来可能有助于观察运动耐力能力的差异。然而,在我们的研究中,注射SFN超过4天并没有改变肌肉纤维的类型和大小(图2A-C)。Nrf2基因的存在与否并不影响纤维的组成和形态。因此,我们认为在目前的力竭测试中,SFN诱导的运动能力提高并不受肌肉成分变化的调节。
我们展示了线粒体生物发生的标记物(图3),以阐明运动能力增加的原因。我们发现四次SFN注射后显著激活AMPKα (图3),与最近的研究一致(32)。然而,我们没有检测其他线粒体生物发生标志物的显著差异,包括四组中mtDNA拷贝数(图3A-C)。此外,我们评估了cAMP的变化(图2H),这被认为是线粒体生物发生的效应(44)。然而,我们没有测量各组之间cAMP的显著差异。Kitaoka等人证明Nrf2基因的存在并不影响线粒体形态的改变(45)。因此,我们假设激活线粒体生物发生信号与通过注射SFN提高运动耐力能力之间的联系较弱,而SFN注射仅进行了四次。
Nrf2 +/+和Nrf2 -/-小鼠的运动能力与一些研究报道的结果不一致(41,46)。我们认识到这种不一致是由于不同的运动耐力测试方案造成的。由于本研究最重要的目的是建立Nrf2 +/+小鼠注射SFN时运动能力和Nrf2 -/-组Nrf2缺失的差异,因此我们更重视运动耐力测试方案。通过试验性实验,我们发现运动试验方案的运动耐力能力差异很大,其中我们选择了最佳试验方案来证明我们的假设。前面提到的研究通过在跑步机上逐渐增加速度和/或倾斜至最大运动水平来测量运动能力。本研究有可能在运动耐力测试中使用厌氧动力而非有氧动力。若以厌氧动力为基础,则小鼠腿的动力无法跟随较高的速度负荷和坡度阻力。因此,遵循测试方案的研究之间结果的不一致是可以想象的。然而,我们认识到保持28米/分钟到力竭是非常适合评估运动耐力能力的。
最近,有关于ROS和肌肉运动适应的重要报道(18,19)。也就是说,抗氧化补充剂可能会妨碍运动训练引起的骨骼肌适应。因此,我们应该谨慎地得出有关长期SFN干预的结论。然而,即使抗氧化剂具有益处,但根据类型、持续时间、频率和剂量,它可能具有毒副作用。因此,我们认为需要更多科学证据证明抗氧化功能。我们不认为SFN仅仅是一种抗氧化剂。当然,很明显,通过SFN干预Nrf2活化的主要作用是间接表达具有抗氧化特性的各种转录因子。然而,难以说明萝卜硫素直接影响其他抗氧化剂以诱导肌肉适应的细胞信号传导。因此,需要更多的实验来回答这个问题,以确定SFN的长期负面影响。
我们的研究结果表明,SFN诱导的Nrf2上调和SFN介导的抗氧化作用可能通过减少力竭运动引起的氧化应激来减轻肌肉疲劳。最终可能会提高运动耐力。 我们相信这项研究为Nrf2在改善运动表现中的作用提供了新的见解。
图1. 实验方案。连续3天,所有小鼠均以5-10米/分的速度低速进行10分的适应性训练。然后,所有小鼠在3天内 SFN或溶媒注射4次(72、48、24和3小时后再进行跑步机测试)。实验当天,所有小鼠均进行该实验方案。运动强度逐渐增加,初始速度为5米/分,然后每3分钟逐渐加速,至最大速度28米/分。保持最大速度,直到小鼠精疲力尽。红色箭头表示SFN或溶媒注射后体内成像的时间进程。结果如图3A[(a)~(b)]所示。
图2. SFN或溶媒注射后在pH 10.8下通过ATP酶染色获得的腓肠肌纤维类型(A)。用ATP酶染色测定腓肠肌中的纤维面积(B)和纤维类型分布(C)。使用间接量热计在每个黑暗和光照循环期间评估每日EE(D)和RQ(E)值。测量腓肠肌ATP(F)、糖原(G)和cAMP(H)水平。
图3. SFN注射效果。SFN或溶媒注射4次,持续3天,并检测小鼠腓肠肌线粒体生物发生标志物。(A)线粒体生物发生标志物的蛋白质表达水平(蛋白质印迹条带)。 将一定量的磷酸化AMPKα标准化为AMPKα蛋白的量。还检测量了线粒体生物发生标志物SirT1和PGC1α的蛋白质含量。载荷体积分别通过肌动蛋白和核纤层蛋白A / C的表达水平标准化。(B)AA线粒体生物发生标志物的mRNA表达。通过实时PCR测定NRF1、TFAM、p53R2、COX IV、SCO1和SCO2的mRNA水平。将值标准化为管家基因GAPDH的水平。(C)腓肠肌mtDNA拷贝数。这个数字影响细胞核和mtDNA比值(mtDNA / nDNA); 括号** P <0.01。
图4. 采用体内成像系统检测SFN诱导的Nrf2- luc活性,并对处于俯卧位的Nrf2和OKD48转基因小鼠(A)进行基因分型。荧光素酶测定的时间过程[(a:基线)?(b:SFN注射后)]如图1所示。使用分光计(B)测量第四次SFN注射后腓肠肌Nrf2-Luc活性。四次SFN注射或溶媒注射后,腓肠肌(C)和比目鱼肌(D)Nrf2靶基因的mRNA水平。括号* P <0.01。
图5. 在力竭跑步试验(A)中,各组小鼠每次跑步的距离以及每次跑步期间的VO2(圆圈)和RQ(三角形)记录了跑步距离中值(B)。各组平均能量底物水平:VO2 (C)、RQ (D)、葡萄糖(E)、FFA (F) ,经过50分钟的跑步机疲劳试验后,各组在同一时间进行比较。括号* P < 0.01。
图6. 腓肠肌氧化、TBARS和GSSG/GSH (A)标记物,以及肌肉损伤、CPK和LDH (B)标记物、肌肉疲劳、血液乳酸(C)标记物。
本文由福山生物整理翻译,转载请注明出处。