






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
英国柳叶刀重磅发布:硒与人类健康(综述)
发表于:2019-02-19 作者:admin 来源:本站 点击量:9105

Margaret P Rayman
(Faculty of Health and Medical Sciences, University of Surrey, Guildford, UK)
摘要:包含在硒蛋白中的硒元素有着广泛且多样的生理功能,包括能影响甲状腺激素活性的抗氧化和抗炎作用。过去10年间,疾病发生与硒蛋白基因的多态性被联系到了一起,从而使得人们重视起了硒蛋白与健康的关系。低硒状态已经被证实与死亡率的风险增加、免疫功能低下,和认知衰退有关。高硒状态或补硒具有抗病毒的效果(这对繁殖而言是必不可少的),并且减少了自身免疫性甲状腺疾病的发生风险。前瞻性研究普遍表明,更高水平的硒对前列腺癌、肺癌、结直肠癌和膀胱癌的发生风险减少有益,但是一些临床试验的结果却喜忧参半。这可能意味着只有在摄入营养物不足的时候补硒才有效。已经有充足的硒摄入的人群继续补硒可能会增加他们患2型糖尿病的风险。需要强调的是,硒保健作用的关键因素是不可分割的U形连接状态:额外硒的摄入也许会对低硒水平的人们有益,而高硒水平的人们补充反而会有不利的影响,因而不应该补硒。
引言
早在十年前就有研究人员认为适当的硒元素水平与健康密切相关。但从那以后,人们对补充硒元素的过分热衷反而导致了不少副作用,这使得人们首次认识到硒元素的毒性[1]。该综述对先前关于硒的研究[2]进行最新阐述,同时着重于临床研究,对目前尚存的争议尤其是硒在癌症和2型糖尿病中的作用进行了讨论。

硒的作用:硒蛋白
在人体内,发挥作用的硒蛋白有25种,它们的共同特点是其活性中心存在硒代半胱氨酸[3]。硒代半胱氨酸的插入由mRNA中UGA密码子确定,但仍需其他因素的相互作用[3–4]。在硒元素摄入不足的情况下,某些硒蛋白的合成(如谷胱甘肽过氧化物酶,GPx4)较其他硒蛋白增多[4]。硒蛋白中很多都是对人体十分重要的酶,从编码硒蛋白基因的单核苷酸多态性(SNP)对疾病和死亡的影响可以看出其对人体健康的重要作用(表1)[21]。
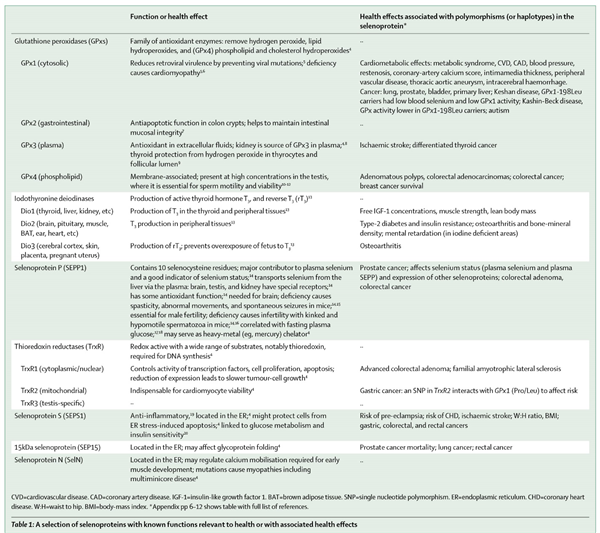
表1 部分已知功能的硒蛋白对健康或健康效应的影响
硒的摄入
相比其他微量营养元素的摄入,硒的摄入量在全球范围内差异很大,从摄入不足导致的硒缺乏病到摄入过量导致的中毒症状(蒜味呼吸、头发指甲脱落、神经系统和皮肤异常、口腔疾病以及麻痹等[22])均有发生。在欧洲,通过饮食摄入的硒元素从7 μg/d到4990 μg/d不等,平均40 μg/d;而在美国,女性摄入的硒元素为93 μg/d,男性为134 μg/d[1,23–24]。硒强化食品可以增加硒元素的摄入量,在美国,约有50%的人在食用硒强化食品[24]。体内硒元素水平可以通过血清或血浆硒元素测定来了解,其因地域和摄入量的不同而有所差异[2]。委内瑞拉、加拿大和美国的硒摄入较高,而日本的硒摄入明显低于欧洲,尤其是东欧。中国同时存在缺硒地区和富硒地区。以前新西兰的硒摄入很低,但自从增加澳大利亚富硒小麦的进口后,硒元素摄入水平明显得到改善[1]。目前推荐的平均硒摄入量男性为60 μg/d,女性为53 μg/d[25]。
硒元素摄入量的变化不但与种植作物土壤的含硒量有关,而且取决于影响硒元素利用的因素(框表),如硒元素的形态、土壤酸碱度、有机物含量以及可以与硒形成化合物的离子含量等[22]。
框表:硒的食物来源
先前的综述已经详细叙述过硒的食物来源了[1,2,24,26]。图1给出了常见食物来源的特征性硒含量[27]。作为主食的谷物中的硒含量,从英国平均0.025-0.033mg/kg(干重)的低硒,到美国高硒地区的30mg/kg不等[26],这是膳食硒摄入量变化的主要原因。虽然巴西坚果是最丰富的硒源,但这是一种不常吃的食物,在任何情况下摄入量变化都很大,所带来的硒摄入量从0.03mg/kg到512mg/kg(鲜重)不等[26]。
在英国,从总膳食研究可以看出不同事物对总膳食硒摄入的贡献。总膳食研究是一项连续的市场调查,调查中,食物代表了英国的平均饮食而被购买、准备、并合并成组进行元素分析(图2)[28]。虽然在其他国家没有同样的数据,但对美国成年人的膳食来源硒的统计结果表明,在美国,面包和谷物对硒的贡献要稍高于英国(37%比26%)[29]。
食物中的硒形式[26]
· 硒代蛋氨酸:蛋氨酸的硒类似物;发现在植物源(尤其是谷物)、酵母硒和其他硒补充剂中。它能非特异性地掺入机体蛋白质中代替蛋氨酸(如白蛋白中的硒代蛋氨酸能让硒在血液中被检测出);因此包含硒代蛋氨酸的补充剂可能含有更多生物硒。
· 硒代半胱氨酸:半胱氨酸的硒类似物;在动物源食物中被发现(来源于动物的硒蛋白)。
· 麦角硒因(Selenoneine) (2-selenyl-Nα,Nα,Nα- trimethyl-l-histidine):一种在金枪鱼和鲭鱼等鱼类中新发现的硒化物形式,也是它们的主要硒化合物;其在鱿鱼,罗非鱼和猪中含量较低[30]。具有很强的清除自由基的活性。
· 甲基硒代半胱氨酸和γ-谷氨酰-硒甲基硒代半胱氨酸:发现在富硒大蒜、洋葱、西兰花等植物中。它能代谢为甲基硒醇,因而被认为具有抗癌作用。
·亚硒酸钠和硒酸盐:硒膳食补充剂的成分;水中偶尔会有硒酸盐,有些硒酸盐也被发现在鱼类和植物源中(比如卷心菜)。
* 这些不同物种的代谢方式[26]和生物利用率[31]被综述在其他地方。
图1:食物源中的不同硒含量
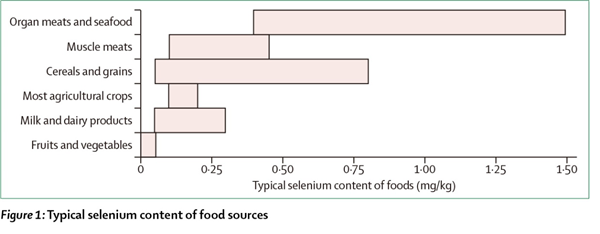
图2:不同食物组对英国硒摄入的贡献
硒对健康的作用
死亡率
至少有3项前瞻性研究显示[32–34],高硒状态与低死亡率有关。在美国第三次全国健康与营养调查行动中,对13 887例参与者长达12年(直到2000年)的随访后发现,硒元素水平与所有原因和癌症造成的死亡率成非线性相关[32]。图3显示了对以上参与者进一步随访到2006年年底的情况。将血浆硒水平提高到约135 μg/L与死亡率降低有关。在一项对法国1389名独立生活的老年人进行的9年纵向流行病学血管老化(EVA)研究发现:低硒(平均87 μg/L)与总死亡率和癌症死亡率的升高有关[33]。巴尔的摩女性健康与衰老研究也发现:低硒是老年女性5年死亡率的独立预测因素[34]。但是,一项对平均年龄为57岁的1103名中国人的15年随访研究,结果并未显示总死亡率与硒元素的基线水平(平均73 μg/L)有相关性[35]。
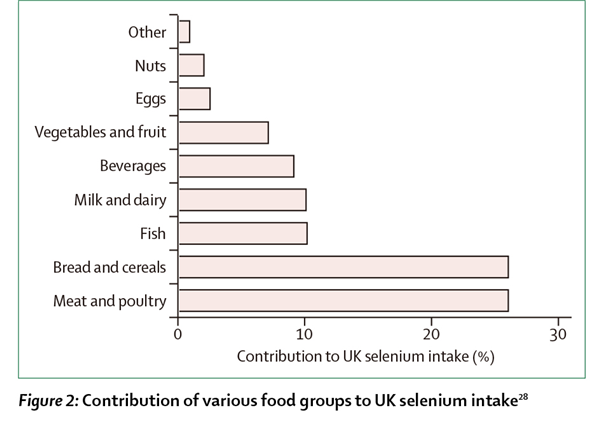
图3美国第三次全国健康和营养调查中(随访18年直到2006年底),成人参与者的血清硒浓度与全因死亡率调整后的风险比
阴影区域显示95%顺式。参考值(危险比1)设置在血清硒分布的第十个百分位(105·8μg/L)。直方图表示研究样本中血清硒浓度的频率分布。
(第三次全国健康和营养调查的死亡率相关文件可见:http://www.cdc.gov/nchs/
data/datalinkage/nh3_mort_analytic_guidelines.pdf)
尽管如此,以上试验并不能完全说明问题,因为营养状态良好的健康人血硒浓度比营养状态差的或处于疾病中的人高[36],这或许提示,在急性反应期,炎性介质会升高而硒元素水平会降低[37–38]。由于肾脏功能的逐渐衰退(肾脏合成血浆GPx3)和亚临床炎症反应的发生,在死亡前几年硒元素水平就开始下降了 [8,38]。
免疫功能
尽管体外研究和动物试验均证实硒对免疫力至关重要[39–41],但人体研究的证据还相对缺乏。即使对于不缺乏硒元素的人群进行补硒,也能发挥促进免疫的作用,包括促进活化T细胞的增殖,提高细胞毒性的淋巴细胞介导的肿瘤细胞毒性以及自然杀伤细胞的活力[2,39,42–45]。在老年人和接受治疗的肿瘤患者中,免疫应答水平通常比较低下。
美国亚利桑那州的一项研究给老年志愿者补充400 μg/d硒元素(以富硒酵母计),结果T细胞计数较安慰剂组提高了27%,主要以CD4+T细胞和细胞毒性自然杀伤细胞的增多为主[42]。一个研究机构给比利时老年居民补充100 μg/d硒元素(以富硒酵母计)连续6个月,结果使其对抗原的反应力达到了正常成人的上限水平[2]。
在头颈部鳞癌患者手术或放疗期间补充200 μg/d硒(亚硒酸钠)可以显著提高患者在治疗期间细胞免疫的反应力[43],而接受安慰剂的患者免疫力有所下降。
只有一项人体研究证实补充硒元素对免疫系统具有功能性疗效[45]。英国缺硒成年人在补充硒元素后,给予口服减毒的脊髓灰质炎病毒,结果发现其病毒清除的速率明显高于安慰剂人群[45]。
上文提到的补硒可以促进CD4+T细胞向Th1细胞分化,而非Th2效应细胞,这与先前研究证实的高硒状态或补硒可以使变应性哮喘患者获益相一致[2,39]。但是,迄今为止所进行的大型随机、双盲、安慰剂对照试验证实[46],补充100 μg/d硒元素(富硒酵母)连续24周并不能使197例英国哮喘患者临床获益,但其中有75%的患者接受吸入类固醇治疗,这有可能抵消潜在的临床获益。
一项对259例HIV-1感染后接受药物治疗患者的病例对照研究发现[47],相比硒浓度较高的患者,血浆硒浓度低于135 μg/L的患者患分枝杆菌病的风险更高(约为前者的3倍),其中3/4为结核。
硒蛋白对活化T细胞的功能至关重要[2]。T细胞尤其对氧化应激敏感,硒蛋白缺乏的T细胞由于无法抑制活性氧产生,因此不能对T细胞受体刺激作出应答[41]。在少数硒代半胱氨酸(Sec)插入序列结合蛋白2(SBP2)杂合缺陷的患者中,其大多数硒蛋白的合成能力都比较低下[48]。这些患者总淋巴细胞计数降低,在接受多克隆刺激后,T细胞的分化能力也显著下降。以上结果证实,硒蛋白在有效免疫应答体系的建立中发挥着重要作用[48]。
白细胞介素2(IL-2)与其受体的相互作用驱使T细胞的增殖[39]。人体研究发现,补充硒元素与淋巴细胞的增殖有关,而这种作用正是通过增加IL-2高亲和受体的表达来实现的[49]。给予富硒食物的小鼠IL-2及其高亲和受体的表达均增加,同时T细胞信号以及在体CD4+T细胞的反应能力均有所增强[39]。高硒饮食将Th1和Th2淋巴细胞之间的平衡向Th1方向推动,从而导致干扰素γ和CD40配体的增加[39],这对基于良好Th1活力的病毒和肿瘤抵抗具有重要作用[39–40,43,45]。
对HIV和其它病毒的作用
硒缺乏(血浆或血清硒浓度≤85 μg/L)与HIV感染者存活时间的减少有关[2]。但与此同时,低硒(不一定缺乏)、低CD4+细胞计数与高病毒载量之间的关系还可归因于严重HIV-1感染者急性反应期血硒浓度的降低[50]。
两项随机对照研究显示,补硒可使HIV感染患者获益[51–52]。在接受药物治疗的美国HIV阳性成人中,补充硒元素200 μg/d可显著减少住院率和感染导致的入院比例[51]。美国的另一项对HIV感染成人的研究发现,即使对抗病毒方案和依从性、HIV感染状态和时间以及丙肝合并感染等进行校正后,较高的血硒浓度仍然预示着病毒载量的降低[52],但也有人对该试验的数据分析方法以及CD4+细胞计数与病毒载量之间的关系等提出质疑[53]。还有一项对坦桑尼亚913名未常规使用药物治疗的HIV感染孕妇(体内硒状态未知)的研究发现,产前和产后每日补充200 μg硒(硒代甲硫氨酸)对HIV-1病毒载量和CD4+细胞的计数无影响,但却减少了6周龄以上儿童的死亡率[53]。
缺硒与其他病毒感染的发生率、毒力和疾病进展有关[2,45]。Beck等人[54]证实,缺硒小鼠体内保护性GPx1活性低下甚至缺失,其可导致RNA病毒的突变,从而产生毒性更强的病毒株。这个发现可以解释导致克山病的柯萨奇病毒突变所致的心肌炎[5–6]。
对大脑的作用
硒对大脑非常重要,当体内硒元素消耗时,大脑会以其他组织的硒损失为代价而保持自身硒元素充足,如果进一步消耗则会造成不可逆性的脑损伤[14]。硒蛋白P(SEPP1)在外周硒元素向大脑的转运中扮演着重要角色,其与脂蛋白受体家族成员apoER2结合介导硒向大脑的转运[14]。无法合成SEEP1的小鼠会出现强直、运动障碍及自发性癫痫发作[14–15]。也有来自于人体研究的证据提示,硒在癫痫、共济协调、帕金森病和认知功能减退中具有一定作用。儿童及成人癫痫患者[55–56]以及热惊厥儿童[57–58]的硒元素水平明显降低。一项小规模研究发现,补硒可以缓解儿童难治性癫痫[2]。
InCHIANTI是一项包含了1012名65岁以上参与者的队列研究[59],结果发现,相对于硒元素水平较高的人,低硒人群的共济协调能力更差,而且研究人员还发现,帕金森病在低硒人群中有增加的趋势。
EPP1具有重要的神经保护作用,其可增强神经元的活力以及预防β样淀粉多肽介导的氧化作用而导致的细胞凋亡[60]。来自人体的研究显示,阿尔茨海默病和痴呆与体内硒状态有关。法国的一项EVA队列研究包含了1166名年龄在60~70岁的参与者[61],结果发现对于低硒人群,认知功能减退的发生风险在4年后明显升高,9年以后认知功能的减退与硒水平下降的程度显著相关[62]。一项对2000名中国农村65岁以上成人的横断面调查研究显示,在80%的人群检测中,较低的指(趾)甲硒水平与较低的认知评分显著有关,且与硒水平存在剂量-效应关系[63]。
然而,在认知功能减退的老年人群中,较低的硒水平在部分程度上可预示肾功能不足导致的GPx3合成减少[8],或者由于炎症因子作用(在急性反应期)导致的硒蛋白合成减少[37]。肾功能减退同时造成同型半胱氨酸(一种已知的痴呆风险因子)在血液中的残留[64]。究竟指甲中的硒元素是否会受到类似的影响尚属未知。
尽管之前有证据[2]表明补硒有益于情绪,但一项对年龄在60~74岁人群的大规模随机安慰剂对照试验显示,以富硒酵母补硒6个月(100 μg/d、200 μg/d或300 μg/d)未见对情绪和生活质量的改善[65]。
生育和繁殖
在男性中,GPx4存在于精子的线粒体鞘中。在精子形成的早期,GPx4作为抗氧化的过氧化物酶起到对精子的保护作用,而在后期,其参与形成中段蛋白的交联,最终变成线粒体鞘的结构成分,对精子的运动能力至关重要[10]。
在一项日本研究中,10%的不育男性和35%的脂质过氧化导致的少弱精子症患者均在精子中发现GPx4缺陷[11]。在这些人群中,精子GPx4表达和精子运动活力消失,其精液中精子数量也减少。但低GPx4水平并不一定由缺硒引起,因为日本人硒摄入量较高[1],而且其在白细胞中的表达水平与正常男性无明显差异[11]。
印度一项研究也发现,不育男性精子中GPx4含量较正常对照组低(93 vs 188 U/mg精子蛋白),其与精子的寿命(r=0.35)、形态完整性(r=0.44)以及运动能力(r=0.45)有关[12]。
睾丸存在一种特异性受体(apoER2)来结合生育所需要的另一种硒蛋白SEPP1[14]。为保持正常浓度和活性的GPx4和SEPP1,每日硒摄入量应该在75 μg左右[66]。在一项随机试验中,每日补充100 μg硒元素可显著改善低硒不育男性精子的运动能力,并使其中11%的男性获得生育功能[2]。但过高的硒摄入(约300 μg/d)会减弱精子的运动能力[67]。
研究发现反复流产的女性往往血硒浓度过低[2,68]。在怀孕期间,硒元素通常会减少,部分原因是由于血容量的增多[68],除此之外,过度的炎症反应(流产的一个可能特性)也会消耗循环中的硒元素[37–38]。
硒摄入与体内硒状态均与子痫前期有关[68–69]。研究显示,发生子痫前期的孕妇血浆和趾甲硒浓度、血浆和胎盘GPx含量以及胎盘硫氧还蛋白还原酶水平均明显低于正常妊娠妇女[69–71]。
英国的一项研究也发现[69],在平均妊娠期为34周的孕妇中,其趾甲(标本取于妊娠前)硒元素浓度的中位数明显低于正常对照组(P<0.001)。趾甲含硒量低的女性在怀孕时发生子痫前期的风险是正常女性的4.4倍[69]。
硒蛋白可通过对抗氧化应激、内质网应激、炎症反应以及保护内皮、控制类花生酸产生、调节血管张力等途径减少子痫前期的发生[68]。系统性炎症反应是子痫前期发生的主要特点之一,硒蛋白(SEPS1)在控制内质网应激和炎症反应中发挥了重要作用[72]。一项挪威的回顾性研究显示[72],发生子痫前期的女性(n=1139)较对照组(n=2269)更有可能携带SEPS1 g.-105G>A 等位基因多态性,从而提示SEPS1在子痫前期发病风险中的作用。
荷兰一项包含1129例妊娠女性的横断面研究发现,早产女性妊娠12周时血浆硒元素水平较正常足月妊娠的女性低[73]。子痫前期同时与硒元素[69]和早产有关,但对以上数据按照子痫前期进行校正之后,血清硒浓度处于最后1/4的女性早产的可能性是其他女性的两倍。但低硒究竟是以上现象的原因还是结果仍有待研究[73]。
甲状腺功能与自身免疫性甲状腺疾病
甲状腺的硒含量高于其他任何组织[13]。硒在甲状腺中具有多种功能,比如非活性甲状腺T4向活性T3的转化需要硒依赖型甲腺原氨酸脱碘酶的参与[13]。一项对体内硒元素处于中低水平的368名甲状腺功能正常的英国老年人进行的随机对照试验,并未显示出补硒对甲状腺功能以及游离(或总)T4向T3的转化率有何影响[74]。
从碘化物到甲状腺球蛋白T3和T4的合成过程中,需要过氧化氢与甲状腺过氧化物酶的参与,而以GPx3形式存在的硒元素可以防止此过程中过氧化氢对甲状腺细胞的损伤[13]。该功能与之前研究得出的“法国女性体内硒元素情况与甲状腺体积、甲状腺组织损伤及甲状腺肿物负相关[75],而挪威人群低硒与甲状腺癌的发生率正相关[76]”这一结论相一致。除此之外,还有不少研究显示,每日补硒80μg或200μg(以亚硒酸钠或硒代甲硫氨酸计)可以对抗桥本甲状腺炎——一种最为常见的自身免疫性甲状腺疾病,以产生补体结合的甲状腺过氧化物酶自身抗体为主要特点[77–78]。一项系统综述的结果显示,补硒可以在3个月时显著降低甲状腺过氧化物酶自身抗体[77]。
患有自身免疫性甲状腺炎的妊娠妇女(甲状腺过氧化物酶自身抗体阳性)更容易发生产后甲状腺功能障碍及永久性甲状腺功能减退。当每日给予200 μg硒代甲硫氨酸时,甲状腺炎症反应明显减轻,而且产后甲状腺功能障碍和永久性甲状腺功能减退的发生率也显著降低[79]。
除此之外,硒也与自身免疫性甲状腺功能亢进有关,比如Graves病。在一项随机对照研究中,给予参与者每日2次亚硒酸钠,每次100 μg,或给予己酮可可碱(600 mg)或安慰剂,连续6个月,结果发现硒元素处理组的生活质量更高,眼部受累更少,Graves眼病进展更慢[80]。
病危
硒元素的摄入与全身炎症反应综合征和脓毒血症有关[81]。有研究发现,存在全身炎症反应综合征或感染性休克患者的血浆硒浓度降低了40%,血浆SEPP1含量降低了70%[82–83]。两项荟萃分析显示,如果给予以上患者高剂量亚硒酸钠,其病死率可出现下降趋势[82]。但治疗需要从大剂量开始(1 mg),因为有研究发现连续低剂量治疗对病死率无影响[82]。
心血管疾病
硒对心血管系统的潜在益处主要来源于其可防止脂质的氧化修饰,抑制血小板聚集及减少炎症反应等一系列证据[2,68,84],以及多态性硒蛋白GPx1、GPx3、Dio2及SEPS1对心脏代谢的影响(表1)。尽管一项对25项观察性试验进行的荟萃分析显示,在低硒人群中,硒元素状态与冠心病呈负相关[85],但关于硒元素补充的一项随机对照试验并未显示其对心脏疾病的预防作用和对死亡终点的影响[85–87]。相比之下,一项对3112名美国成人的研究没有显示趾甲硒浓度与亚临床动脉粥样硬化(颈动脉中层增厚、冠状动脉硬化评分)之间具有相关性[88]。
有些横断面研究发现高硒状态与血浆胆固醇的升高有关[84]。英国一项对501名低硒老年人群的随机试验显示[84],在对所有参与者进行为期6个月的补硒干预(100 μg/d和200 μg/d,以富硒酵母计)后,其血浆总胆固醇与非HDL胆固醇均显著降低,但补充300 μg/d反而使HDL胆固醇升高。随着硒剂量的升高,总胆固醇与HDL胆固醇的比值随之降低,提示补硒对减少心血管疾病的风险有益,至少在以上研究人群中可以得出此结论。其他两项小规模研究并未得出以上结论[89–90]。
研究对象体内硒元素水平的不同或许可以对以上差异做出解释。除了特定的血硒浓度(该浓度下硒蛋白具有最佳活性)外,过高的血硒浓度对减少心血管死亡率无益,也有研究显示其为U型相关[32]。比如,在英国PRECISE试验中,在参与者硒水平的基线浓度仅有一半人的GPx1处于最佳活性,在这种情况下,补硒的潜在获益就可显现[84]。相比之下,在美国的一项研究[86–87]中,大多数硒蛋白在研究对象硒元素的基线水平就已经处于最佳活性[66,91–92],因此单纯补充硒元素无效,除非提高硒蛋白水平。一项对636名低硒疑似冠心病人群(平均血浆硒浓度74 μg/L)的队列研究结果显示,在为期4.7年的随访后,基线红细胞GPx1活性是后续心血管事件的强烈预测因子[93]。
癌症
前瞻性研究发现硒可以减少肺癌[94]、膀胱癌[95]、结肠直肠癌[96]、肝癌[97]、食管癌[35]、贲门癌[35]、甲状腺癌[76]以及前列腺癌的风险[21,96,98–100]。表2总结了补充硒元素对肺癌、膀胱癌和前列腺癌的影响。

表2 硒与癌症风险前瞻性研究的荟萃分析(按组织分类)
早期的一项大规模病例对照研究显示,基于食物的硒摄入量与肺癌的发生呈负相关[103],但两项后续研究并未发现硒摄入[101]和硒状态[102]与肺癌的发生风险存在相关性。一项对非洲裔美国人的多种族队列研究发现,硒元素处于较高水平的人患前列腺癌的风险比硒水平处于最低1/4的人低41%[108],但除了这项研究以外,尚无其他研究证明硒与前列腺癌风险的相关性[104–107]。然而,EPIC-海德堡队列研究显示,对于血清硒浓度处于第3个四分位数(并非第4个,≥95.0 μg/L)的人群,其前列腺癌的发生风险比处于第1个四分位数的人低,这在高级别疾病中表现得更加明显[107]。
一项综述[96]对近期关于前列腺癌的3项研究[106–108]进行了总结,发现硒元素与晚期疾病的保护相关,而在局部或低级别疾病中并无此作用,且以上相关性在吸烟人群中表现得更加强烈。
以硒作为单一营养元素进行干预比较少见。一项系统综述和荟萃分析[109]对来自中国江苏省启东市的3项关于补充抗氧化剂对胃肠道肿瘤预防作用的研究进行了总结,启东人口大约有15%乙肝病毒阳性,尽管结果显示补充抗氧化剂使肝癌的发生率降低了50%,但该试验的质量遭到诸多质疑。
营养对肿瘤的预防(NPC)试验募集了来自美国的1312名之前患有非黑素瘤性皮肤癌病史的志愿者[110],结果显示每日补充200 μg硒(以富硒酵母计),平均持续4.5年,对非黑素瘤性皮肤癌的主要预后没有影响,但经过6.4年的随访,补硒明显减少了癌症病死率(50%)和癌症的总发病率(37%),使前列腺癌、结肠直肠癌及肺癌的发病率分别减少了67%、58%和46%。但在后来的分析中,排除血浆硒水平处于基线水平1/3以下(<106 μg/L)的患者后[112],仅癌症总发病率的减少(25%)和前列腺癌的减少(52%)具有统计学意义[111]。对于硒浓度处于最高1/3水平(>122-123 μg/L)的患者,其总癌症和前列腺癌的患病风险有所升高(分别为20%和14%),但差异无统计学意义。除此以外,硒浓度处于最高2/3的人群鳞癌的发生率明显升高,但硒浓度处于最低1/3的人(血浆硒含量>106 μg/L)该风险并没有显著降低[111,113–114]。对于基线硒水平高于122 μg/L的参与者,补硒对预防癌症不但无益,反而有可能增加罹患癌症的风险。
一项关于硒和维生素E的研究(SELECT)募集了35 533名美国男性,目的在于探讨以上两种元素对前列腺癌发病风险的影响。对以上参与者经过5.5年的随访发现,补硒200 μg/d(硒代甲硫氨酸)不能减少基线硒水平在136 μg/L的人群局部前列腺癌的发病风险[87]。但该研究没有包括血浆硒水平<106 μg/L的参与者,之前研究显示对于血硒处于该水平的人补硒可减少前列腺癌的发病风险[111,113]。在SELECT研究中,硒元素基线水平的四分位数间距为122.4~151.8 μg/L,根据NPC研究,对于硒元素在该范围内的人群补硒没有使患癌症的风险明显增高。事实上,研究人员还报道了补硒的潜在毒副作用,比如脱发和皮炎[87]。在SELECT研究中,补硒的形式是硒代甲硫氨酸而非富硒酵母,后者30%~40%的硒不是硒代甲硫氨酸[25]。
SELECT试验没有对以下问题进行解释:①硒对晚期疾病风险的影响,不同的研究证实硒对晚期疾病的影响更大[96,100,115],因为在SELECT试验中仅有1%的病例为非局部疾病;②硒对前列腺癌死亡率的影响,因为仅有1例患者死于前列腺癌[87];③硒对近期吸烟者的影响,有研究显示硒对吸烟者的保护作用更强[96],但在SELECT研究中,吸烟者仅占7.5%;④硒对低硒状态人群的影响,在该研究中仅小部分参与者属于低硒人群。在NPC研究中,给予补硒之前,有1/3参与者的SEPP1或GPx浓度不在最佳水平,而在SELECT研究中并不存在此情况,相反,大多数参与者硒蛋白浓度和活性的基线水平都相当高[116]。
以下因素或许可以解释研究之间的差异:第一,硒对预防前列腺癌的进展可能更为重要,因此其对晚期疾病的影响可能比早期更大[23,96];第二,硒元素只有在特定浓度范围内才能发挥其减少癌症患病风险的作用,既不能太低也不能太高,否则都不能使GPx和SEPP1保持最佳活性,然而SELECT研究中的参与者硒浓度过高,英国营养素与癌症前瞻性研究(EPIC)中纳入的参与者体内硒浓度又过低[21,87,106,117]。与其他营养素类似,不少基于人体的研究发现,营养素的摄入或其状态与癌症的预防呈U形相关[1,96,111];第三,硒元素的摄入或其状态与遗传背景之间可能存在重要的相互作用,硒蛋白基因的单核苷酸多态性(SNP)可影响硒蛋白的合成、其在血浆中的浓度和活性以及疾病的发生风险[21]。例如,GPx1基因的SNP可以影响前列腺癌、肺癌、乳腺癌及膀胱癌的发病风险;SEPP1、GPx4和SEPS1的SNP可以影响结肠直肠癌的发病风险;GPx3的SNP与甲状腺癌的发病风险有关;SEP15的变异与肺癌、直肠癌及前列腺癌的存活期有关;GPx4的SNP可以影响乳腺癌的存活期(表1)。除此之外,以上这些硒蛋白的SNP(及其作用通路相关的SNP)可与体内硒状态相互作用,因此存在特定基因变异的人或许可从额外的硒元素补充中获得更多的益处[115,118]。
2型糖尿病
硒与糖代谢之间的相关性目前仍存争议[119]。有三项病例对照研究显示[120–122],高硒状态可以减少糖尿病的发生,前瞻性EVA研究通过对男性患者9年的随访后发现[123],较高的血浆硒浓度可以改善高糖血症。
相比之下,美国大型全民营养与健康调查研究则发现,高硒与糖尿病发病风险的升高有关[124–125]。无独有偶,法国SUVIMAX试验显示[126]血浆硒元素水平与空腹血糖正相关。
两项将2型糖尿病作为次要结局的随机对照试验得出的结论也不一致。SELECT研究经过5.5年的随访发现,给予35 533名参与者每日补硒200 μg,对2型糖尿病的发病风险无影响[87]。但一项对1312名美国东南部参与者进行的事后分析显示,经过7.7年的随访,每日补硒200 μg(以富硒酵母计)会使患2型糖尿病的风险升高[127]。暴露-反应关系梯度显示,对于体内硒浓度处于基线水平前1/3(>121.6 μg/L)的参与者,补硒更容易使其2型糖尿病的发病风险升高[127]。
那么如何解释不同试验得出的不同结论?在上述病例对照研究中,2型糖尿病患者体内硒元素较低,这可能是疾病及其相关的炎症反应造成的影响。比如,全身炎症反应产生的细胞因子可抑制SEPP1的表达,同时消耗体内硒元素[37–38,83]。但这种关系并不能完全解释EVA研究中硒对高血糖症的改善[123]。氧化应激可以诱发胰岛素抵抗,抗氧化治疗对胰岛素抵抗的改善也间接证明了以上观点[128]。EVA研究包含了体内硒浓度处于维持抗氧化剂GPx最佳活性所需硒浓度之上和之下的患者,结果显示,硒浓度处于上1/3的参与者氧化应激损伤较下1/3者小(中位血浆硒浓度为104 μg/L vs 71 μg/L)[123],因此前者不易发生胰岛素抵抗。但为何没有在女性患者中发现相似的作用还有待研究。
对于高硒摄入可能增加2型糖尿病的风险则归因于高硒对胰岛素信号通路的影响。胰岛素与其受体的结合激发了一系列级联反应,其中伴随过氧化氢这一第二信使的参与[119]。高活性GPx1可以清除过氧化氢,从而干预胰岛素信号通路。例如,过表达GPx1的转基因小鼠会发生胰岛素抵抗、高血糖症、高胰岛素血症以及肥胖[129]。在妊娠妇女中也观察到了红细胞GPx1活性与轻度胰岛素抵抗之间较强的相关性[130]。相比之下,敲除GPx1基因可以改善小鼠体内胰岛素诱导的糖摄取,同时改善胰岛素抵抗[119]。然而,GPx1并不是唯一与以上作用相关的硒蛋白,因为在远低于增加2型糖尿病风险的硒浓度下,GPx就已经达到最大活性,而且已有证据表明Dio2的多态性可以影响胰岛素抵抗和2型糖尿病的发生风险表1)。事实上,硒蛋白合成能力低下的人群(主要为SBP2杂合缺陷)对胰岛素的敏感性更强[48]。
另一个与糖尿病风险有关的硒蛋白是SEPP1,其达到最大血浆浓度比GPx1需要更多的硒摄入[66]。SEPP1发挥负性胰岛素调节作用:其可在体外抑制胰岛素诱导的活性氧产生,而且可通过钝化AMP活化的蛋白酶(一种存在于胰岛B细胞中的胰岛素合成和分泌的正性调节剂)来影响胰岛素抵抗[17]。日本一项临床试验显示,SEPP1浓度与糖化血红蛋白A1c和空腹血糖有关,而且在2型糖尿病人群中其浓度升高[17]。在韩国2型糖尿病患者及处于糖尿病前期的人群中,其体内SEPP1浓度明显高于糖耐量正常的人群,而且下降速度缓慢[18]。除此之外,SEPP1在肥胖和超重的人群中更高[18],提示碳水化合物代谢失调可以通过激活PGC1α而使SEPP1表达上调,其中PGC1α是一种转录因子的共激活因子,在肝脏糖异生和SEPP1的生物合成中具有重要作用[119]。
但为何在NPC研究中补硒使2型糖尿病的发病风险升高,而在SELECT试验中却无此作用?其实小规模的NPC研究相比大规模的SELECT研究具有更多发现糖尿病的机会。SELECT参与者硒元素的基线水平较NPC研究的参与者高(平均血清硒136 μg/L vs 平均血浆硒114 μg/L[87,127]),该浓度的硒元素已经使得硒蛋白的表达和活性在额外补充硒元素之前就已经达到风险阈值。SELECT试验中每1000(人·年)安慰剂组的例数比NPC试验高(14.1 vs 8.4)。
在动物模型中,GPx1及其他应激相关硒蛋白(通过食物中硒元素的摄入量调节)的低表达和高表达均与胰岛素抵抗和高糖血症有关[131]。2型糖尿病与硒蛋白之间的U型相关可以在一定程度上解释矛盾的试验结果。
硒对健康的影响
不同地区人群体内硒元素水平具有较大差异,其与硒摄入量一致[1–2]。图4显示了英国[132]和美国[133]人群血浆硒元素分布的不同。图中的黑色点状垂直线(位于122 μg/L)代表NPC试验[110–111,114,127]中是否影响癌症、非黑素瘤性皮肤癌、2型糖尿病等疾病风险的血浆硒元素基线浓度分界线。该图显示:血浆或血清硒浓度≥122 μg/L的人群(在美国该人群所占比例相当大)无需补硒。
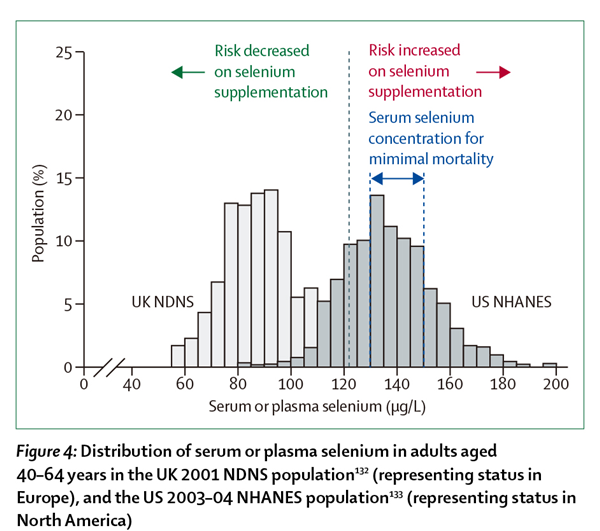
图4 英国2001年NDNS人群(代表欧洲)和美国2003-2004年间NHANES人群[133](代表北美)中的40-64岁成年人血清和血浆硒的分布[132]。
柱状图的高度代表具有相应的血清或血浆硒含量的人群的加权百分比。122μg/L的黑色垂直虚线表示血浆硒的基线浓度。这些血浆硒描述了在癌症营养预防临床试验中,每天补充200微克的硒后,非黑素瘤皮肤癌和2型糖尿病从低到高的风险变化[110,111,114,127]。
蓝色虚线显示了在第三NHANS人群中,与最低死亡率相关的血浆硒浓度分布[32]。直方图由Guallar和Yiyi Zhang构建。NDNS即全国饮食与营养调查(National Diet and Nutrition Survey)。NHANES即为全国健康和营养调查(National Health and Nutrition Examination Survey.)
反之同样成立,对于血浆或血清硒浓度<122 μg/L的人群,补硒不仅没有额外风险,而且对健康有益,将硒浓度提高至130~150 μg/L都是可以接受的,因为此浓度与最低死亡率相关(图3)[32]。
结论及对未来研究的建议
硒对人体健康的影响具有多样性和复杂性[1],因此未来的研究应着眼于如何优化益处同时减少风险。对于低硒或相对低硒人群的试验研究还有待进一步开展。既然硒蛋白的多态性对体内硒状态、疾病及其预后都有影响(表1),那么以后的研究就应该选取不同基因型的参与者为研究对象,进行更细致的研究。今后的研究应该首先侧重于揭示硒与2型糖尿病高发之间的潜在关系。
对于低硒人群,额外补充硒元素(如强化食品)将有较大益处,但高硒人群则无需额外补充。这一U型关系与Paracelsus早在1567年得出的规律一致。
原文:Rayman M P. Selenium and human health.[J]. Lancet, 2012, 379(9822):1256-1268.
References
1 Rayman MP. Food-chain selenium and human health: emphasis on intake. Br J Nutr 2008; 100: 254–68.
2 Rayman MP. The importance of selenium to human health. Lancet 2000; 356: 233–41.
3 Kryukov GV, Castellano S, Novoselov SV, et al. Characterization of mammalian selenoproteomes. Science 2003; 300: 1439–43.
4 Reeves MA, Hoff mann PR. The human selenoproteome: recent insights into functions and regulation. Cell Mol Life Sci 2009; 66: 2457–78.
5 Beck MA, Levander OA, Handy J. Selenium defi ciency and viral infection. J Nutr 2003; 133 (suppl 1): 1463S–67S.
6 Lei C, Niu X, Wei J, Zhu J, Zhu Y. Interaction of glutathione peroxidase-1 and selenium in endemic dilated cardiomyopathy. Clin Chim Acta 2009; 399: 102–08.
7 Florian S, Krehl S, Loewinger M, et al. Loss of GPx2 increases apoptosis, mitosis, and GPx1 expression in the intestine of mice. Free Radic Biol Med 2010; 49: 1694–702.
8 Schiavon R, Guidi GC, Biasioli S, De Fanti E, Targa L. Plasma glutathione peroxidase activity as an index of renal function. Eur J Clin Chem Clin Biochem 1994; 32: 759–65.
9 Schmutzler C, Mentrup B, Schomburg L, Hoang-Vu C, Herzog V, K?hrle J. Selenoproteins of the thyroid gland: expression, localization and possible function of glutathione peroxidase 3. Biol Chem 2007; 388: 1053–59.
10 Ursini F, Heim S, Kiess M, et al. Dual function of the selenoprotein PHGPx during sperm maturation. Science 1999; 285: 1393–96.
11 Imai H, Suzuki K, Ishizaka K, et al. Failure of the expression of phospholipid hydroperoxide glutathione peroxidase in the spermatozoa of human infertile males. Biol Reprod 2001; 64: 674–83.
12 Foresta C, Flohé L, Garolla A, Roveri A, Ursini F, Maiorino M. Male fertility is linked to the selenoprotein phospholipid hydroperoxide glutathione peroxidase. Biol Reprod 2002; 67: 967–71.
13 Schomburg L, K?hrle J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol Nutr Food Res 2008; 52: 1235–46.
14 Burk RF, Hill KE. Selenoprotein P-expression, functions, and roles in mammals. Biochim Biophys Acta 2009; 1790: 1441–47.
15 Schweizer U, Br?uer AU, K?hrle J, Nitsch R, Savaskan NE. Selenium and brain function: a poorly recognized liaison. Brain Res Brain Res Rev 2004; 45: 164–78.
16 Olson GE, Winfrey VP, Nagdas SK, Hill KE, Burk RF. Selenoprotein P is required for mouse sperm development. Biol Reprod 2005; 73: 201–11.
17 Misu H, Takamura T, Takayama H, et al. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab 2010; 12: 483–95.
18 Yang SJ, Hwang SY, Choi HY, et al. Serum selenoprotein P levels in patients with type 2 diabetes and prediabetes: implications for insulin resistance, infl ammation, and atherosclerosis. J Clin Endocrinol Metab 2011; 96: E1325–29.
19 Curran JE, Jowett JB, Elliott KS, et al. Genetic variation in selenoprotein S influences infl ammatory response. Nat Genet 2005; 37: 1234–41.
20 Gao Y, Walder K, Sunderland T, et al. Elevation in Tanis expression alters glucose metabolism and insulin sensitivity in H4IIE cells. Diabetes 2003; 52: 929–34.
21 Rayman MP. Selenoproteins and human health: insights from epidemiological data. Biochim Biophys Acta 2009; 1790: 1533–40.
22 Johnson CC, Fordyce FM, Rayman MP. Symposium on ‘Geographical and geological infl uences on nutrition’: factors controlling the distribution of selenium in the environment and their impact on health and nutrition. Proc Nutr Soc 2010; 69: 119–32.
23 Rayman MP. Selenium in cancer prevention: a review of the evidence and mechanism of action. Proc Nutr Soc 2005; 64: 527–42.
24 Fairweather-Tait SJ, Bao Y, Broadley MR, et al. Selenium in human health and disease. Antioxid Redox Signal 2011; 14: 1337–83.
25 Rayman MP. The use of high-selenium yeast to raise selenium status: how does it measure up? Br J Nutr 2004; 92: 557–73.
26 Rayman MP, Infante HG, Sargent M. Food-chain selenium and human health: spotlight on speciation. Br J Nutr 2008; 100: 238–53.
27 WHO. Selenium. A report of the International Programme on Chemical Safety. Environmental Health Criteria number 58. Geneva: World Health Organization, 1987.
28 Food Standards Agency. Survey on measurement of the concentrations of metals and other elements from the 2006 UK total diet study. 2009. http://www.food.gov.uk/multimedia/pdfs/ fsis0109metals.pdf (accessed Feb 14, 2011).
29 Cotton PA, Subar AF, Friday JE, Cook A. Dietary sources of nutrients among US adults, 1994 to 1996. J Am Diet Assoc 2004; 104: 921–30.
30 Yamashita Y, Yamashita M. Identifi cation of a novel seleniumcontaining compound, selenoneine, as the predominant chemical form of organic selenium in the blood of bluefin tuna. J Biol Chem 2010; 285: 18134–38.
31 Fairweather-Tait SJ, Collings R, Hurst R. Selenium bioavailability: current knowledge and future research requirements. Am J Clin Nutr 2010; 91: 1484S–91S.
32 Bleys J, Navas-Acien A, Guallar E. Serum selenium levels and all-cause, cancer, and cardiovascular mortality among US adults. Arch Intern Med 2008; 168: 404–10.
33 Akbaraly NT, Arnaud J, Hininger-Favier I, Gourlet V, Roussel AM, Berr C. Selenium and mortality in the elderly: results from the EVA study. Clin Chem 2005; 51: 2117–23.
34 Ray AL, Semba RD, Walston J, et al. Low serum selenium and total carotenoids predict mortality among older women living in the community: the women’s health and aging studies. J Nutr 2006; 136: 172–76.
35 Wei WQ, Abnet CC, Qiao YL, et al. Prospective study of serum selenium concentrations and esophageal and gastric cardia cancer, heart disease, stroke, and total death. Am J Clin Nutr 2004; 79: 80–85.
36 Bates CJ, Thane CW, Prentice A, Delves HT. Selenium status and its correlates in a British national diet and nutrition survey: people aged 65 years and over. J Trace Elem Med Biol 2002; 6: 1–8.
37 Hesse-Bahr K, Dreher I, Kohrle J. The infl uence of the cytokines Il-1beta and INFgamma on the expression of selenoproteins in the human hepatocarcinoma cell line HepG2. Biofactors 2000; 11: 83–85.
38 Nichol C, Herdman J, Sattar N, et al. Changes in the concentrations of plasma selenium and selenoproteins after minor elective surgery: further evidence for a negative acute phase response? Clin Chem 1998; 44: 1764–66.
39 Hoffmann FW, Hashimoto AC, Shafer LA, Dow S, Berry MJ, Hoffmann PR. Dietary selenium modulates activation and diff erentiation of CD4+ T cells in mice through a mechanism involving cellular free thiols. J Nutr 2010; 140: 1155–61.
40 Hoffmann PR. Mechanisms by which selenium influences immune responses. Arch Immunol Ther Exp (Warsz) 2007; 55: 289–97.
41 Carlson BA, Yoo MH, Shrimali RK, et al. Role of seleniumcontaining proteins in T-cell and macrophage function. Proc Nutr Soc 2010; 69: 300–10.
42 Wood SM, Beckham C, Yosioka A, Darban H, Watson RR. β-Carotene and selenium supplementation enhances immune response in aged humans. Integr Med 2000; 2: 85–92.
43 Kiremidjian-Schumacher L, Roy M, Glickman R, et al. Selenium and immunocompetence in patients with head and neck cancer. Biol Trace Elem Res 2000; 73: 97–111.
44 Hawkes WC, Kelley DS, Taylor PC. The eff ects of dietary selenium on the immune system in healthy men. Biol Trace Elem Res 2001; 81: 189–213.
45 Broome CS, McArdle F, Kyle JA, et al. An increase in selenium intake improves immune function and poliovirus handling in adults with marginal selenium status. Am J Clin Nutr 2004; 80: 154–62.
46 Shaheen SO, Newson RB, Rayman MP, et al. Randomised, double blind, placebo-controlled trial of selenium supplementation in adult asthma. Thorax 2007; 62: 483–90.
47 Shor-Posner G, Miguez MJ, Pineda LM, et al. Impact of selenium status on the pathogenesis of mycobacterial disease in HIV-1-infected drug users during the era of highly active antiretroviral therapy. J Acquir Immune Defic Syndr 2002; 29: 169–73.
48 Schoenmakers E, Agostini M, Mitchell C, et al. Mutations in the selenocysteine insertion sequence-binding protein 2 gene lead to a multisystem selenoprotein defi ciency disorder in humans. J Clin Invest 2010; 120: 4220–35.
49 Roy M, Kiremidjian-Schumacher L, Wishe HI, Cohen MW, Stotzky G. Supplementation with selenium and human immune cell functions. I. Eff ect on lymphocyte proliferation and interleukin 2 receptor expression. Biol Trace Elem Res 1994; 41: 103–14.
50 Drain PK, Baeten JM, Overbaugh J, et al. Low serum albumin and the acute phase response predict low serum selenium in HIV-1 infected women. BMC Infect Dis 2006; 6: 85.
51 Burbano X, Miguez-Burbano MJ, McCollister K, et al. Impact of a selenium chemoprevention clinical trial on hospital admissions of HIV-infected participants. HIV Clin Trials 2002; 3: 483–91.
52 Hurwitz BE, Klaus JR, Llabre MM, et al. Suppression of human immunodefi ciency virus type 1 viral load with selenium supplementation: a randomized controlled trial. Arch Intern Med 2007; 167: 148–54.
53 Kupka R, Mugusi F, Aboud S, et al. Randomized, double-blind, placebo-controlled trial of selenium supplements among HIV-infected pregnant women in Tanzania: eff ects on maternal and child outcomes. Am J Clin Nutr 2008; 87: 1802–08.
54 Beck MA, Handy J, Levander OA. Host nutritional status: the neglected virulence factor. Trends Microbiol 2004; 12: 417–23.
55 Ashrafi MR, Shabanian R, Abbaskhanian A, et al. Selenium and intractable epilepsy: is there any correlation? Pediatr Neurol 2007; 36: 25–29.
56 Ashrafi MR, Shams S, Nouri M, et al. A probable causative factor for an old problem: selenium and glutathione peroxidase appear to play important roles in epilepsy pathogenesis. Epilepsia 2007; 48: 1750–55.
57 Amiri M, Farzin L, Moassesi ME, Sajadi F. Serum trace element levels in febrile convulsion. Biol Trace Elem Res 2010; 135: 38–44.
58 Mahyar A, Ayazi P, Fallahi M, Javadi A. Correlation between serum selenium level and febrile seizures. Pediatr Neurol 2010; 43: 331–34.
59 Shahar A, Patel KV, Semba RD, et al. Plasma selenium is positively related to performance in neurological tasks assessing coordination and motor speed. Mov Disord 2010; 25: 1909–15.
60 Takemoto AS, Berry MJ, Bellinger FP. Role of selenoprotein P in Alzheimer’s disease. Ethn Dis 2010; 20 (suppl 1): S1-92–95.
61 Berr C, Balansard B, Arnaud J, Roussel AM, Alperovitch A. Cognitive decline is associated with systemic oxidative stress: the EVA study. Etude du Vieillissement Arteriel. J Am Geriatr Soc 2000; 48: 1285–91.
62 Akbaraly NT, Hininger-Favier I, Carriere I, et al. Plasma selenium over time and cognitive decline in the elderly. Epidemiology 2007; 18: 52–58.
63 Gao S, Jin Y, Hall KS, et al. Selenium level and cognitive function in rural elderly Chinese. Am J Epidemiol 2007; 165: 955–65.
64 Ravaglia G, Forti P, Maioli F, et al. Homocysteine and cognitive function in healthy elderly community dwellers in Italy. Am J Clin Nutr 2003; 77: 668–73.
65 Rayman M, Thompson A, Warren-Perry M, et al. Impact of selenium on mood and quality of life: a randomized, controlled trial. Biol Psychiatry 2006; 59: 147–54.
66 Xia Y, Hill KE, Li P, et al. Optimization of selenoprotein P and other plasma selenium biomarkers for the assessment of the selenium nutritional requirement: a placebo-controlled, double-blind study of selenomethionine supplementation in selenium-deficient Chinese subjects. Am J Clin Nutr 2010; 92: 525–31.
67 Hawkes WC, Turek PJ. Eff ects of dietary selenium on sperm motility in healthy men. J Androl 2001; 22: 764–72.
68 Rayman MP. Selenium and adverse health conditions of human pregnancy. In: Hatfi eld DL, Berry MJ, Gladyshev VN, eds. Selenium: its molecular biology and role in human health, 3rd edn. New York: Springer Science and Business Media, 2011: 531–46.
69 Rayman MP, Bode P, Redman CW. Low selenium status is associated with the occurrence of the pregnancy disease preeclampsia in women from the United Kingdom. Am J Obstet Gynecol 2003; 189: 1343–49.
70 Vanderlelie J, Venardos K, Clifton VL, Gude NM, Clarke FM, Perkins AV. Increased biological oxidation and reduced anti-oxidant enzyme activity in pre-eclamptic placentae. Placenta 2005; 26: 53–58.
71 Mistry HD, Wilson V, Ramsay MM, Symonds ME, Broughton Pipkin F. Reduced selenium concentrations and glutathione peeroxidase activity in preeclamptic pregnancies. Hypertension 2008; 52: 881–88.
72 Moses EK, Johnson MP, T?mmerdal L, et al. Genetic association of preeclampsia to the infl ammatory response gene SEPS1. Am J Obstet Gynecol 2008; 198: 336 e1–5.
73 Rayman MP, Wijnen H, Vader H, Kooistra L, Pop V. Maternal selenium levels during early gestation and risk for preterm birth. CMAJ 2011; 183: 549–55.
74 Rayman MP, Thompson AJ, Bekaert B, et al. Randomized controlled trial of the eff ect of selenium supplementation on thyroid function in the elderly in the United Kingdom. Am J Clin Nutr 2008; 87: 370–78.
75 Derumeaux H, Valeix P, Castetbon K, et al. Association of selenium with thyroid volume and echostructure in 35- to 60-year-old French adults. Eur J Endocrinol 2003; 148: 309–15.
76 Glattre E, Thomassen Y, Thoresen SO, et al. Prediagnostic serum selenium in a case-control study of thyroid cancer. Int J Epidemiol 1989; 18: 45–49.
77 Toulis KA, Anastasilakis AD, Tzellos TG, Goulis DG, Kouvelas D. Selenium supplementation in the treatment of Hashimoto’s thyroiditis: a systematic review and a meta-analysis. Thyroid 2010; 20: 1163–73.
78 Nacamulli D, Mian C, Petricca D, et al. Infl uence of physiological dietary selenium supplementation on the natural course of autoimmune thyroiditis. Clin Endocrinol (Oxf) 2010; 73: 535–39.
79 Negro R, Greco G, Mangieri T, Pezzarossa A, Dazzi D, Hassan H. The infl uence of selenium supplementation on postpartum thyroid status in pregnant women with thyroid peroxidase autoantibodies. J Clin Endocrinol Metab 2007; 92: 1263–68.
80 Marcocci C, Kahaly GJ, Krassas GE, et al, European Group on Graves’ Orbitopathy. Selenium and the course of mild Graves’ orbitopathy. N Engl J Med 2011; 364: 1920–31.
81 Rayman MP. The argument for increasing selenium intake. Proc Nutr Soc 2002; 61: 203–15.
82 Forceville X. Eff ects of high doses of selenium, as sodium selenite, in septic shock patients a placebo-controlled, randomized, double-blind, multi-center phase II study – selenium and sepsis. J Trace Elem Med Biol 2007; 21 (suppl 1): 62–65.
83 Renko K, Hofmann PJ, Stoedter M, et al. Down-regulation of the hepatic selenoprotein biosynthesis machinery impairs selenium metabolism during the acute phase response in mice. FASEB J 2009; 23: 1758–65.
84 Rayman MP, Stranges S, Griffi n BA, Pastor-Barriuso R, Guallar E. Eff ect of supplementation with high-selenium yeast on plasma lipids: a randomized, controlled trial. Ann Intern Med 2011; 154: 656–65.
85 Flores-Mateo G, Navas-Acien A, Pastor-Barriuso R, Guallar E. Selenium and coronary heart disease: a meta-analysis. Am J Clin Nutr 2006; 84: 762–73.
86 Stranges S, Marshall JR, Trevisan M, et al. Eff ects of selenium supplementation on cardiovascular disease incidence and mortality: secondary analyses in a randomized clinical trial. Am J Epidemiol 2006; 163: 694–99.
87 Lippman SM, Klein EA, Goodman PJ, et al. Eff ect of selenium and vitamin E on risk of prostate cancer and other cancers: the Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2009; 301: 39–51.
88 Xun P, Liu K, Morris JS, Daviglus ML, He K. Longitudinal association between toenail selenium levels and measures of subclinical atherosclerosis: the CARDIA trace element study. Atherosclerosis 2010; 210: 662–67.
89 Luoma PV, Sotaniemi EA, Korpela H, Kumpulainen J. Serum selenium, glutathione peroxidase activity and high-density lipoprotein cholesterol–eff ect of selenium supplementation. Res Commun Chem Pathol Pharmacol 1984; 46: 469–72.
90 Yu SY, Mao BL, Xiao P, et al. Intervention trial with selenium for the prevention of lung cancer among tin miners in Yunnan, China. A pilot study. Biol Trace Elem Res 1990; 24: 105–08.
91 Duffield AJ, Thomson CD, Hill KE, Williams S. An estimation of selenium requirements for New Zealanders. Am J Clin Nutr 1999; 70: 896–903.
92 Hurst R, Armah CN, Dainty JR, et al. Establishing optimal selenium status: results of a randomized, double-blind,placebo-controlled trial. Am J Clin Nutr 2010; 91: 923–31.
93 Blankenberg S, Rupprecht HJ, Bickel C, et al. Glutathione peroxidase 1 activity and cardiovascular events in patients with coronary artery disease. N Engl J Med 2003; 349: 1605–13.
94 Zhuo H, Smith AH, Steinmaus C. Selenium and lung cancer: a quantitative analysis of heterogeneity in the current epidemiological literature. Cancer Epidemiol Biomarkers Prev 2004; 13: 771–78.
95 Amaral AF, Cantor KP, Silverman DT, Malats N. Selenium and bladder cancer risk: a meta-analysis. Cancer Epidemiol Biomarkers Prev 2010; 19: 2407–15.
96 Peters U, Takata Y. Selenium and the prevention of prostate and colorectal cancer. Mol Nutr Food Res 2008; 52: 1261–72.
97 Yu MW, Horng IS, Hsu KH, Chiang YC, Liaw YF, Chen CJ. Plasma selenium levels and risk of hepatocellular carcinoma among men with chronic hepatitis virus infection. Am J Epidemiol 1999; 150: 367–74.
98 Etminan M, FitzGerald JM, Gleave M, Chambers K. Intake of selenium in the prevention of prostate cancer: a systematic review and meta-analysis. Cancer Causes Control 2005; 16: 1125–31.
99 Brinkman M, Reulen RC, Kellen E, Buntinx F, Zeegers MP. Are men with low selenium levels at increased risk of prostate cancer? Eur J Cancer 2006; 42: 2463–71.
100 Rayman MP. Selenium. In: Milner JA, Romagnolo DF, eds. Bioactive compounds and cancer. New York: Humana Press/ Springer, 2010: 411–48.
101 Mahabir S, Forman MR, Dong YQ, Park Y, Hollenbeck A, Schatzkin A. Mineral intake and lung cancer risk in the NIH-American Association of Retired Persons Diet and Health study. Cancer Epidemiol Biomarkers Prev 2010; 19: 1976–83.
102 Epplein M, Franke AA, Cooney RV, et al. Association of plasma micronutrient levels and urinary isoprostane with risk of lung cancer: the multiethnic cohort study. Cancer Epidemiol Biomarkers Prev 2009; 18: 1962–70.
103 Mahabir S, Spitz MR, Barrera SL, Beaver SH, Etzel C, Forman MR. Dietary zinc, copper and selenium, and risk of lung cancer. Int J Cancer 2007; 120: 1108–15.
104 Peters U, Foster CB, Chatterjee N, et al. Serum selenium and risk of prostate cancer-a nested case-control study. Am J Clin Nutr 2007; 85: 209–17.
105 Peters U, Littman AJ, Kristal AR, Patterson RE, Potter JD, White E. Vitamin E and selenium supplementation and risk of prostate cancer in the Vitamins and lifestyle (VITAL) study cohort. Cancer Causes Control 2008; 19: 75–87.
106 Allen NE, Appleby PN, Roddam AW, et al. European Prospective Investigation into Cancer and Nutrition. Plasma selenium concentration and prostate cancer risk: results from the European Prospective Investigation into Cancer and Nutrition (EPIC). Am J Clin Nutr 2008; 88: 1567–75.
107 Steinbrecher A, Méplan C, Hesketh J, et al. Eff ects of selenium status and polymorphisms in selenoprotein genes on prostate cancer risk in a prospective study of European men. Cancer Epidemiol Biomarkers Prev 2010; 19: 2958–68.
108 Gill JK, Franke AA, Steven Morris J, et al. Association of selenium, tocopherols, carotenoids, retinol, and 15-isoprostane F(2t) in serum or urine with prostate cancer risk: the multiethnic cohort. Cancer Causes Control 2009; 20: 1161–71.
109 Bjelakovic G, Nikolova D, Simonetti RG, Gluud C. Antioxidant supplements for prevention of gastrointestinal cancers: a systematic review and meta-analysis. Lancet 2004; 364: 1219–28.
110 Clark LC, Combs GF Jr, Turnbull BW, et al. Eff ects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional Prevention of Cancer Study Group. JAMA 1996; 276: 1957–63.
111 Duffield-Lillico AJ, Reid ME, Turnbull BW, et al. Baseline characteristics and the eff ect of selenium supplementation on cancer incidence in a randomized clinical trial: a summary report of the Nutritional Prevention of Cancer Trial. Cancer Epidemiol Biomarkers Prev 2002; 11: 630–39.
112 Reid ME, Duffi eld-Lillico AJ, Garland L, Turnbull BW, Clark LC, Marshall JR. Selenium supplementation and lung cancer incidence: an update of the nutritional prevention of cancer trial. Cancer Epidemiol Biomarkers Prev 2002; 11: 1285–91.
113 Duffield-Lillico AJ, Dalkin BL, Reid ME, et al. Nutritional Prevention of Cancer Study Group. Selenium supplementation, baseline plasma selenium status and incidence of prostate cancer: an analysis of the complete treatment period of the Nutritional Prevention of Cancer Trial. BJU Int 2003; 91: 608–12.
114 Duffield-Lillico AJ, Slate EH, Reid ME, et al. Nutritional Prevention of Cancer Study Group. Selenium supplementation and secondary prevention of nonmelanoma skin cancer in a randomized trial. J Natl Cancer Inst 2003; 95: 1477–81.
115 Cooper ML, Adami H-O, Gr?nberg H, Wiklund F, Green FR, Rayman MP. Interaction between SNPs in selenoprotein P and mitochondrial superoxide dismutase determines prostate cancer risk. Cancer Res 2008; 68: 10171–77.
116 Rayman MP, Combs GF Jr, Waters DJ. Selenium and vitamin E supplementation for cancer prevention. JAMA 2009; 301: 1876.
117 Bekaert B, Rayman MP. Plasma selenium concentration and prostate cancer risk. Am J Clin Nutr 2009; 89: 1276–77.
118 Penney KL, Schumacher FR, Li H, et al. A large prospective study of SEP15 genetic variation, interaction with plasma selenium levels, and prostate cancer risk and survival. Cancer Prev Res (Phila) 2010; 3: 604–10.
119 Steinbrenner H, Speckmann B, Pinto A, Sies H. High selenium intake and increased diabetes risk: experimental evidence for interplay between selenium and carbohydrate metabolism. J Clin Biochem Nutr 2011; 48: 40–45.
120 Navarro-Alarcon M, López-G de la Serrana H, Perez-Valero V, Lopez Martinez C. Serum and urine selenium concentrations as indicators of body status in patients with diabetes mellitus. Sci Total Environ 1999; 228: 79–85.
121 Kljai K, Runje R. Selenium and glycogen levels in diabetic patients. Biol Trace Elem Res 2001; 83: 223–29.
122 Rajpathak S, Rimm E, Morris JS, Hu F. Toenail selenium and cardiovascular disease in men with diabetes. J Am Coll Nutr 2005; 24: 250–56.
123 Akbaraly TN, Arnaud J, Rayman MP, et al. Plasma selenium and risk of dysglycemia in an elderly French population: results from the prospective Epidemiology of Vascular Ageing Study. Nutr Metab (Lond) 2010; 7: 21–27.
124 Bleys J, Navas-Acien A, Guallar E. Serum selenium and diabetes in U.S. adults. Diabetes Care 2007; 30: 829–34.
125 Laclaustra M, Navas-Acien A, Stranges S, Ordovas JM, Guallar E. Serum selenium concentrations and diabetes in U.S. adults: National Health and Nutrition Examination Survey (NHANES) 2003–2004. Environ Health Perspect 2009; 117: 1409–13.
126 Czernichow S, Couthouis A, Bertrais S, et al. Antioxidant supplementation does not aff ect fasting plasma glucose in the Supplementation with Antioxidant Vitamins and Minerals (SU. VI.MAX) study in France: association with dietary intake and plasma concentrations. Am J Clin Nutr 2006; 84: 395–99.
127 Stranges S, Marshall JR, Natarajan R, et al. Eff ects of long-term selenium supplementation on the incidence of type 2 diabetes: a randomized trial. Ann Intern Med 2007; 147: 217–23.
128 Houstis N, Rosen ED, Lander ES. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature 2006; 440: 944–48.
129 McClung JP, Roneker CA, Mu W, et al. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc Natl Acad Sci USA 2004; 101: 8852–57.
130 Chen X, Scholl TO, Leskiw MJ, Donaldson MR, Stein TP. Association of glutathione peroxidase activity with insulin resistance and dietary fat intake during normal pregnancy. J Clin Endocrinol Metab 2003; 88: 5963–68.
131 Labunskyy VM, Lee BC, Handy DE, Loscalzo J, Hatfield DL, Gladyshev VN. Both maximal expression of selenoproteins and selenoprotein defi ciency can promote development of type 2 diabetes-like phenotype in mice. Antioxid Redox Signal 2011; 14: 2327–36.
132 Ruston D, Hoare S, Henderson L, et al. The National Diet and Nutrition Survey: adults aged 19 to 64 years. Nutritional status (anthropometry and blood analytes), blood pressure and physical activity (vol 4). London: Stationery Office, 2004.
133 National Center for Health Statistics. National Health and Nutrition Examination Survey. 2003–04. US Department of Health and Human Services, Centers for Disease Control. http://www.cdc.gov/ nchs/nhanes.htm (accessed March 13, 2011).
本文源于祖超译,如有侵权,请联系删除。