






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
miR-24-1-5p的上调参与了黑树莓花色苷对结直肠癌的化学预防作用
发表于:2019-05-13 作者:admin 来源:本站 点击量:9857
摘要
作为重要的表观遗传调控因子,microRNA通过触发靶mRNA的降解和/或通过抑制其翻译来调节蛋白质的表达。已报道在包括结直肠癌的一些癌症中存在microRNA表达失调。在这项研究中,microRNA阵列差异分析显示,在用黑树莓花色苷喂养9周的偶氮甲烷/葡聚糖硫酸钠诱导的小鼠结肠组织中,miR-24-1-5p的表达显著增强。在人类结直肠癌细胞中,miR-24-1-5p的过度表达显著抑制了β-连环蛋白的表达,同时降低了细胞的增殖、迁移和存活。此外,mir-24-1-5p可以靶向β-连环蛋白并触发β-连环蛋白及其下游靶基因的负调控环。β-连环蛋白信号对人结直肠癌的形成和发展至关重要。因此,目前的研究发现,miR-24-1-5p是β-连环蛋白的有效调节因子,这可能为β-连环蛋白信号驱动的结直肠癌提供一种新的化学预防和治疗策略。
结直肠癌(CRC)是全球第三大恶性肿瘤,约占所有癌症发病率和死亡率的10 %。尽管过去20年里人们致力于CRC诊断和治疗策略,但仍有五分之一的患者被诊断为CRC时已进入晚期,通常采用手术和辅助化疗相结合的方法进行治疗(3-5)。大多数患者发生化疗耐药,癌症可能再次发生在这些患者身上并最终转移(6)。最近一些研究已经报道了microRNA(miRNA)表达与CRC(7-9)进展的显著关联。因此,在RNA网络分析的基础上,寻找一种可用于预防或治疗结直肠癌的新靶点是十分必要的。
miRNA是一种长度为18-22 nt的内源性非编码小RNA,通过结合mRNA的3′-UTR(未翻译区域)来干扰其翻译,从而调节基因表达,直接降解结合的mRNA(10,11)。在许多人类癌症中,miRNA的异常表达可激活肿瘤抑制基因或癌基因。在结直肠癌中,有几个miRNA被鉴定为肿瘤发育的必要条件。Michael等人(12)首次报道了人结直肠癌组织和正常结肠直肠粘膜中miRNA的表达差异,共检测到28个差异表达的miRNA,包括miR-320、miR-321、miR-200c、miR-223和miR-145。Monzo等人(13)发现,miR-17-5p可以靶向E2F转录因子1,该转录因子存在于人类结肠早期胚胎发育和肿瘤发生时期并调节细胞增殖。其他研究者报道,miR-34a直接并负向控制p53的一系列下游靶基因,形成miR-34a-p53的正反馈环(14)。Bandres等人(15)发现,miRNA-451可以调节迁移抑制因子(MIF)的表达,从而抑制CRC细胞的增殖,提高肿瘤对放疗的敏感性。此外,miRNA-320a和miR-139可分别靶向β-连环蛋白和RAP1B(RAS癌基因家族成员),抑制人结肠癌细胞的增殖(16,17)。2001年,Lagos Quintana等人(18)在研究非脊柱动物和脊柱动物时首次发现了miR-24。人的miR-24由miR-24-1和miR-24-2组成,分别位于9号和19号染色体上,成熟序列包括miR-24-3p、miR-24-1-5p和miR-24-2-5p(19)。有报道称,miR-24-1在数种癌症组织中表达异常(20-26)。然而,miR-24-1-5p在CRC中的功能尚不清楚。
黑树莓(BRB)属于莓类中的Rubus occidentalis科,富含黄酮类和酚酸(27,28)。BRB对结直肠癌的化学预防作用已被本研究组和其他研究组(29-32)的研究所证实。然而,很少有研究关注miRNA与BRB在结直肠癌中的化学预防作用及其潜在机制的相关性。
在本研究中,我们假设miR-24-1-5p在结直肠癌的发生和发展中起重要作用,也参与了BRB对结直肠癌的化学预防作用。为了验证这一假设,我们在小鼠上进行了一项体内研究,并对人类细胞系进行了数项体外研究,发现miR-24-1-5p可以作为肿瘤抑制因子发挥作用,其靶向数个CRC相关通路中的β-连环蛋白相关癌基因。我们目前的发现可能有助于发展新的诊断和治疗策略,用以预防和治疗结直肠癌。
方法
化合物与试剂
氧化偶氮甲烷(AOM,A5486)购自Sigma Aldrich公司,葡聚糖硫酸钠(DSS,0216011091)购自MP公司。建丰自然产品技术有限公司提供BRB花色苷(纯度>90%)。BRB花色苷由三种主要花色苷——矢车菊素葡萄糖苷、矢车菊素木糖基芦丁糖苷和矢车菊素芸香糖苷组成,含量分别为2.63、0.73和16.91 mg/g。矢车菊素桑布双糖苷的含量不太丰富,我们最近发表的论文(33)详细介绍了这一点。β-连环蛋白、细胞周期蛋白D1、c-Myc和细胞周期蛋白依赖性激酶4(CDK4)从细胞信号技术公司获得,β-肌动蛋白、E-钙粘蛋白、糖原合酶激酶3β(GSK3-β)、磷酸糖原合酶激酶3β(p-GSK3-β)、B细胞淋巴瘤-2(BCL-2)和血凝素从Sangon Biotech有限公司获得。分泌型卷曲相关蛋白2(SFRP2)购自Abcam公司。从Biorbyt Explore生物试剂公司中购得分泌型卷曲相关蛋白5(SFRP5)。免疫印迹的二级抗体为辣根过氧化物酶偶联的山羊抗兔和山羊抗鼠IgG,从ComWin生物科技有限公司获得。
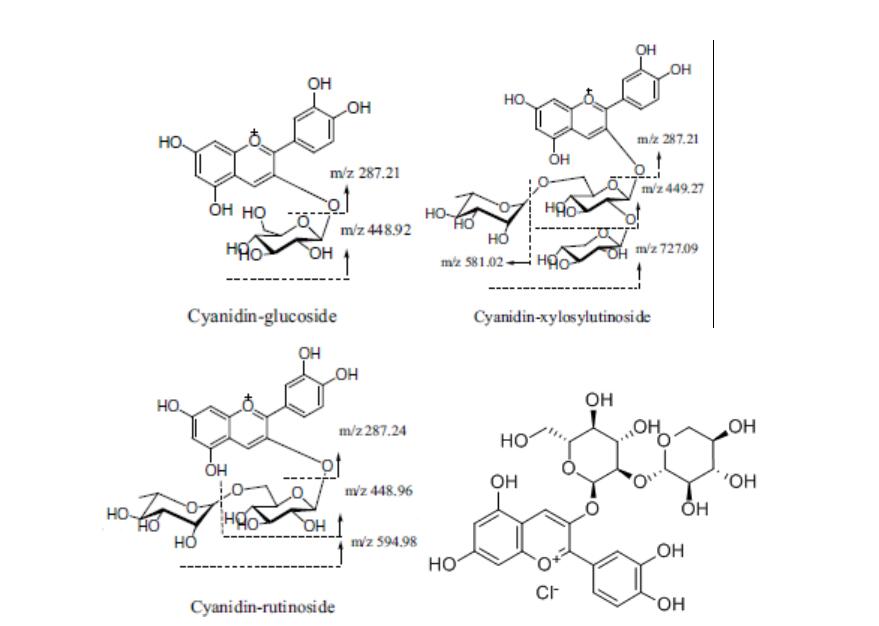
(译者注:化学结构式从左至右分别为矢车菊素葡萄糖苷、矢车菊素木糖基芦丁糖苷、矢车菊素芸香糖苷与矢车菊素桑布双糖苷)
动物护理和实验方案
5周大的雄性C57BL/6J小鼠(18-20g,本溪长生实验动物技术有限公司)给予随意饮水,维持在12小时光暗循环下。这些动物被关在笼子里(每笼5只动物),房间温度设置为21±2.0°C,湿度控制50±5%。所有动物实验方案均获辽宁中医药大学伦理委员会(中国,沈阳)批准。
结肠炎诱导的结直肠癌小鼠模型的建立及黑树莓花色苷的补充
在1周适应期后,小鼠单次腹腔注射AOM(10 mg/kg体重)。1周后,在饮用水中给予2% DSS持续1周,接着给予正常水持续2周,该循环再重复2次。
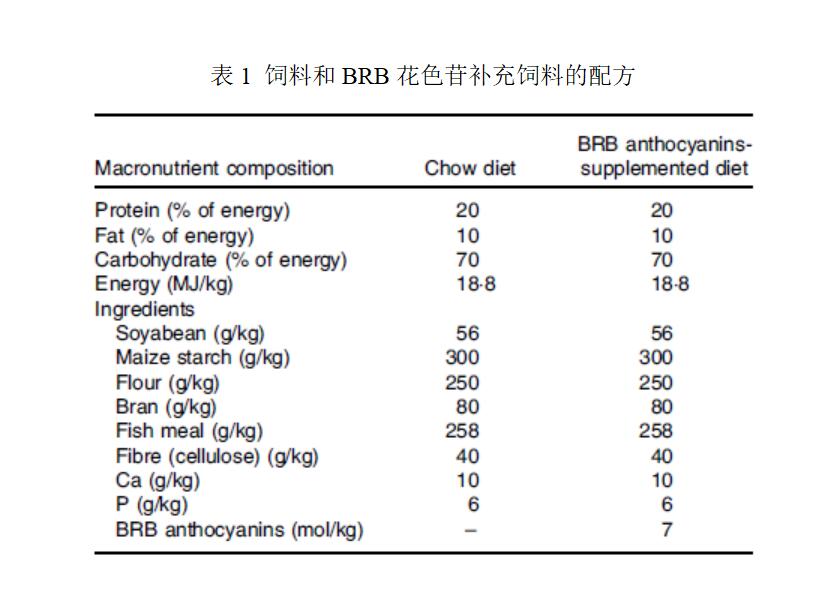
小鼠分为三组:一组健康对照组和两组AOM/DSS处理组。每组10只。健康对照组的小鼠没有接受任何处理,喂以饲料超过9周。对于两个AOM/DSS处理组,一组只用饲料喂养,而另一组则用含有7.0μmol/g BRB花色苷的饲料(10%BRB)喂养。添加饲料中BRB花色苷浓度相当于10%冻干BRB粉中的花色苷含量,这是基于前人研究(32,34)选定的。饲料和BRB花色苷补充饲料的配方组成如表1所示。实验中的所有饲料使用前储存于-20 ℃。
在第9周结束后,小鼠吸入二氧化碳后颈椎脱臼处死。结肠组织立即冷冻在液氮中,然后储存在-80℃下准备进行microRNA阵列分析。
表1 饲料和BRB花色苷补充饲料的配方
MicroRNA微阵列表达谱与数据分析
miRNA表达谱由Agilent Mouse miRNA(8×60K,设计ID:070155;Agilent Technologies公司)生成。总RNA用NanoDrop ND-2000(Thermo Scientific)定量。用Agilent Bioanalyzer 2100(Agilent Technologies公司)评估RNA完整性。总RNA去磷酸化、变性后,用花青苷-3-胞苷三磷酸标记。纯化后,标记的RNA与微阵列杂交。清洗后,使用Agilent Scanner G2505C(Agilent Technologies公司)对阵列进行扫描。阵列图像用Feature Extraction软件(版本10.7.1.1;Agilent Technologies公司)获取。使用GeneSpring软件(版本13.1;Agilent Technologies公司)分析原始数据。
RNA提取与定量实时PCR
总RNA使用Trizol试剂盒(Comwin Biotech有限公司)从细胞中分离,并使用miRNA cDNA合成试剂盒(ComWin Biotech有限公司)通过逆转录产生第一链互补DNA(cDNA)编码的带poly A尾的miRNA。实时PCR(RT-PCR)采用SYBR Green方法以特异性引物进行。根据制造商的说明,RT-PCR在Applied Biosystems 7500实时PCR系统上进行扩增,并用microRNA实时分析试剂盒(ComWin Biotech有限公司)进行分析。引物序列为:mmu-miR-24-1-5p:正链5′-AGTGCCTACTGAGCTGATATCAGT-3′;hsa-miR-24-1-5p:正链5′-GTGCCTACTGAGCTGATATCAGT-3′;β-连环蛋白:正链 5′-GCTGGTGACAGGGAAGACAT-3′,反链5′-AGGCAAACCGTGAAAAGATG-3′;细胞周期蛋白D1:正链5′-AACTACCTGGACCGCTTCCT-3′,反链 5′-CCACTTGAGCTTGTTCACCA-3′;c-Myc:正链5′-TTCGGGTAGTGGAAAACCAG-3′,反链5′-CAGCAGCTCGAATTTCTTCC-3′;CDK4:正链 5′-ATTGGTGTCGGTGCCTATG-3′,反链5′-AACTGTGCTGATGGGAAGG-3′。这些基因的表达水平按照2–ΔΔCT方法以内部对照(U6:正链 5′-ATTGGAACGATACAGAGAAGATT-3′,反链5′-GGAACGCTTCACGAATTTG-3′或β-肌动蛋白:正链5′-AGGCAAACCGTGAAAAGATG-3′,反链 5′-AGGCAAACCGTGAAAAGATG-3′)正常化。
(译者注:正链(forward strand),与mRNA序列相同的DNA单链,另一条则为反链(reverse strand);互补DNA链中携带编码蛋白质信息的链称为正义链(sense strand),又称编码链(coding strand),另一条称为反义链(antisense strand),又称模板链(template strand)。)
质粒构建和转染
miR-24-1-5p的成熟序列为UGCCUACUGAGCUGAUAUCAGU。人pri miR-24-1-5p序列构建使用如下引物:miR-24-1-5p正链, 5′-CCGCTCGAGCAACAGGGTTTTCCAAGTCTAC-3′;miR-24-1-5p反链,5′-GGAAGATCTTCACCTAAGTCGGAAATCATGTGGTA-3′。PCR产物用XhoI和BglII 这一对内切酶消化,插入到XhoI-BglII切开的pcDNA3-血凝素(质粒载体)中。然后用lipofectamine 2000(Invitrogen)将HCT-116和Caco-2细胞与人pri-miR-24-1-5p一起转染。用空白质粒转染的细胞作为阴性对照。构建的人pri-miR-24-1-5p经测序验证。
(备注:成熟的miRNAs是由较长的初级转录物经过一系列核酸酶的剪切加工而产生的,初级转录物称为pri-miRNA。pri-miRNA长度从几百到几千个碱基不等,带有5‘帽子和3’poly A尾巴,以及1到数个发夹径环结构。pri-miRNA经剪切产生约70个碱基的miRNA前体,即pre-miRNA。pre-miRNA为单一发夹结构,5‘带有磷酸基团,3’有两个突出碱基,并带有3‘羟基。pre-miRNA经进一步剪切,形成长度约为22个碱基的单链成熟miRNA。)
细胞培养
人结直肠癌细胞系HCT-116(CCL-247;美国模式培养物集存库(ATCC,American type culture collection))和Caco-2(北京癌症研究所细胞库)分别在Dulbecco改进的Eagle's培养基和α-最低基础培养基(Hyclone,Thermo Fisher Scientific公司)中培养。这两种培养基中都添加10%(v/v)胎牛血清(大连美伦生物)、10000 U/ml青霉素G、10 mg/ml链霉素,在37℃的5% CO2湿润空气中孵育。
伤口愈合试验
伤口愈合试验采用汇合度为70%的HCT-116和Caco-2细胞。首先,用无菌移液管尖端在细胞单层上划一条直线。用PBS洗涤细胞,然后加入新鲜培养基。然后用miR-24-1-5p转染,24小时后,丢弃培养基,用显微照片计算划痕中剩余的间隙。
细胞增殖和群落形成分析
用 3-(4,5-二甲基-2-噻唑)-2,5-二苯基四氮唑溴盐(MTT)法测定细胞活力。简单来说,在miR-24-1-5p转染24小时后,用0.5 mg/ml MTT试剂培养Caco-2和HCT-116细胞4小时。然后去除培养基,在每个孔中加入100μl二甲亚砜,以溶解甲赞晶体。在490nm处测量板的吸光度。为了进一步研究miR-24-1-5p在癌细胞存活中的作用,将细胞以200/孔接种在六孔板中,培养2周进行菌落形成分析。出现的细胞群落首先用PBS洗涤,然后用甲醇固定,再用0.5%的吉姆萨染液(Giemsa,天青-伊红)染色。用倒置显微镜计数每孔中出现的群落数。
Transwell迁移分析
将Caco-2和HCT-116细胞接种在Transwell孔的上室,在下室中加入无血清培养基。24小时后,用棉签去除膜顶部的细胞,然后用4%多聚甲醛固定Transwell孔插入膜。迁移到膜底的细胞用4’,6-二脒基-2-苯基吲哚(北京鼎国昌盛生物技术有限公司)染色。从每个插入膜中随机选择10个域,在20倍放大的显微镜下进行捕获。使用Image Pro Plus 6.0对细胞数进行量化。为了计算迁移指数,将每个处理组捕获的细胞数归一化为对照组细胞数。
microRNA的靶向预测
利用Miranda 3.3a版的算法和网站工具(http://www.microrna.org/microrna/home.do)对miR-24-1-5p的靶基因进行了预测,并给出了默认参数和截止值(评分S≥140,能量E≤-7.0)。
蛋白质印迹
细胞在含1 mmol/L苯甲磺酰氟(北京鼎国昌盛生物技术有限公司)的冷冻放射免疫沉淀分析缓冲液中置于冰上30分钟。在4℃下14000 g离心15 分钟,收集上清液,用10%凝胶SDS-PAGE法测定含有40μg蛋白的上清液。电泳后,将凝胶中的蛋白质转移到硝酸纤维素膜((Sangon Biotech有限公司)上,用5%牛血清白蛋白V(Roche)在Tris缓冲盐水和吐温20缓冲液中封闭2小时,然后在室温下与合适的一级抗体孵育3小时,然后上相应的二级抗体。用增强化学发光技术(Amersham Life Science)检测印迹上的信号。
免疫组织化学染色
收集AOM/DSS诱导小鼠肠道组织,无论是否含有BRB花色苷,固定在10%的福尔马林缓冲液中,并如前所述嵌入石蜡中(35)。组织切片(4μm厚)用抗E-钙粘蛋白抗体(Santa Cruz生物技术)进行免疫组化染色。流程与先前报道类似(35)。简言之,切片脱蜡、复水,3% H2O2淬灭,10%的正常血清封闭,然后用β-连环蛋白抗体在4℃孵育过夜,接着用二级抗体和亲和素-生物素复合物(ABC试剂盒;Vector Laboratory)孵育。用3’,5’-二氨基联苯胺进行染色,再用苏木精进行轻微复染。彩色载玻片的图像由Aperio Image系统采集。
统计方法
采用SPSS 19.0统计软件对数据进行分析。为了比较组内和组间的差异,采用t检验和方差分析,然后进行Fisher最小显著性差异分析。在P<0.05水平上考虑统计学意义。
结论
组织microRNA微阵列分析
为了阐明miRNA介导的结直肠癌肿瘤发生通路,识别异常表达的miRNA是第一步。基于这一点,我们进行了组织miRNA微阵列分析,以获得AOM/DSS诱导小鼠(无论是否补充BRB花色苷)结肠组织中总miRNA表达特征。两组结果的比较表明,通过给予BRB花色苷,数个miRNA的调节有差异(图1(a))。组织阵列的层次聚类分析结果如图1(b)所示。AOM/DSS诱导组表达低水平的miR-24-1-5p,但用BRB花色苷喂养9周后,miR-24-1-5p水平显著上调,提示miR-24-1-5p可能对肿瘤形成有抑制作用。这也证实了BRB花色苷的化学预防功能。
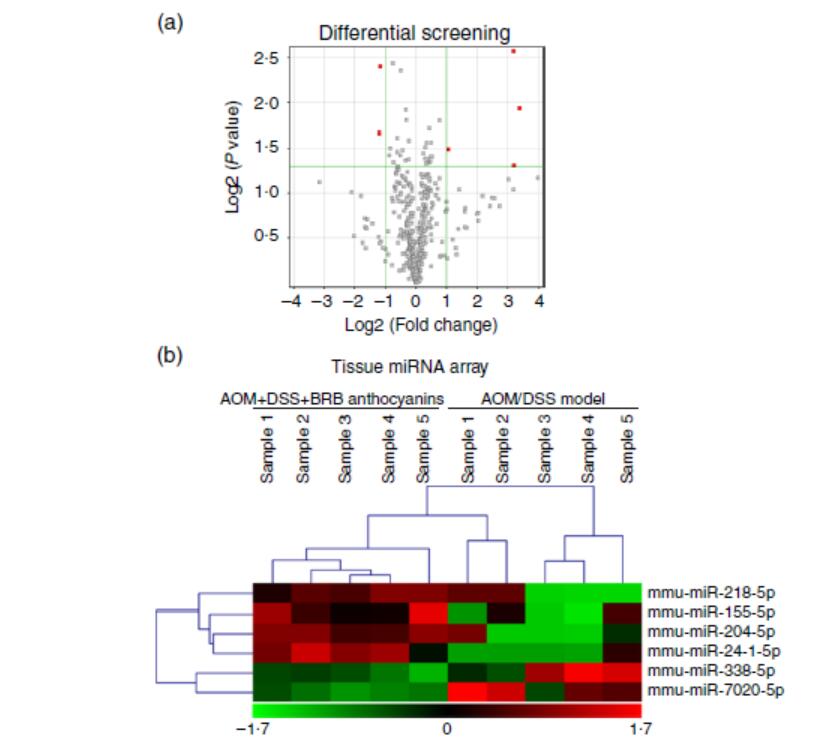
图1 mircoRNA(miRNA)在小鼠结肠组织中的表达差异。(a)上调的miRNA用红色表示。(b)组织miRNA阵列。绿色-下调;红色-上调(n=5)。AOM,偶氮甲烷;DSS,葡聚糖硫酸钠;BRB,黑树莓。
miR-24-1-5p在偶氮甲烷/葡聚糖硫酸钠诱导的小鼠结肠组织中表达下调,在用黑树莓花色苷处理的人结直肠癌细胞系中表达上调
为了进一步评估小鼠模型微阵列分析结果,采用定量RT-PCR(qRT-PCR)分析了有与无BRB花色苷给药的AOM/DSS诱导组结肠组织中的miR-24-1-5p表达。qRT-PCR获得的miR-24-1-5p的表达模式与微阵列数据一致(图2(a))。在用不同剂量的BRB花色苷处理后,测定了几种人结直肠癌细胞系(Caco-2、HCT-116、Lovo、HT29和SW480)中miR-24-1-5p的表达。BRB花色苷导致miR-24-1-5p水平升高。在BRB花色苷处理的人类肿瘤细胞中,miR-24-1-5p的表达模式与用BRB花色苷补充饮食喂养的小鼠结肠组织获得的miRNA微阵列和qRT-PCR数据一致(图2(b))。
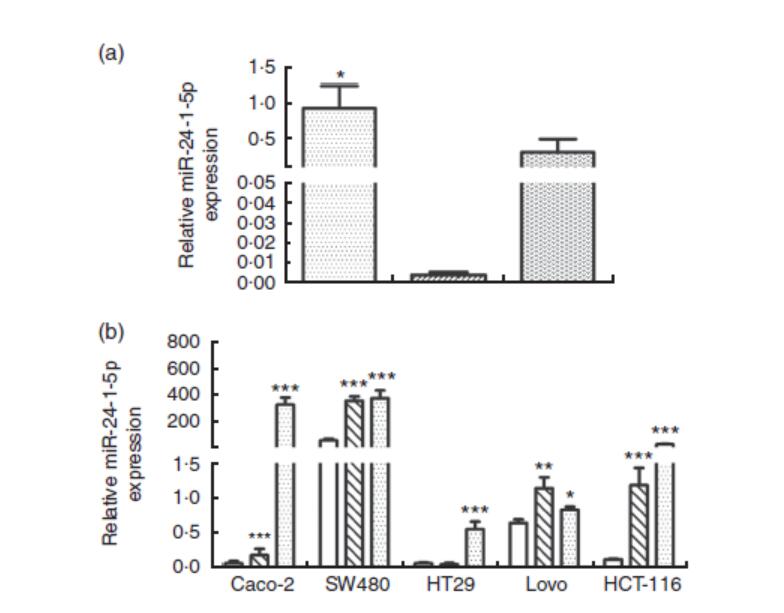
图2 定量RT-PCR(qRT-PCR)检测结肠组织和结直肠癌(CRC)细胞中miR-24-1-5p的表达。(a)qRT-PCR分析添加或不添加黑树莓(BRB)花色苷的偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导小鼠结肠组织中的miR-24-1-5p水平。(b)在不同的CRC细胞系中,相对的miR-24-1-5p表达。单因素方差分析用于比较不同组的人结直肠癌细胞系或小鼠结肠组织。值为平均值(n=5),其标准误差由竖线表示。与溶剂对照组比较,平均值有显著性差异:*P<0.05,**P<0.01,***P<0.001。(a) ,对照;
,对照; ,AOM+DSS模型;
,AOM+DSS模型; ,AOM+DSS+BRB花色苷。(b)
,AOM+DSS+BRB花色苷。(b) ,对照;25μg/ml BRB花色苷;
,对照;25μg/ml BRB花色苷; ,50μg/ml BRB花色苷。
,50μg/ml BRB花色苷。
黑树莓花色苷可降低偶氮甲烷/葡聚糖硫酸钠诱导小鼠的β-连环蛋白含量。
为了研究β-连环蛋白在BRB花色苷原位调节的AOM/DSS诱导小鼠中的作用,我们进行了免疫组化染色。与不给予BRB花色苷饮食的AOM/DSS诱导小鼠相比,给予BRB花色苷饮食的AOM/DSS诱导小鼠小肠的β-连环蛋白水平显著降低(图3)。
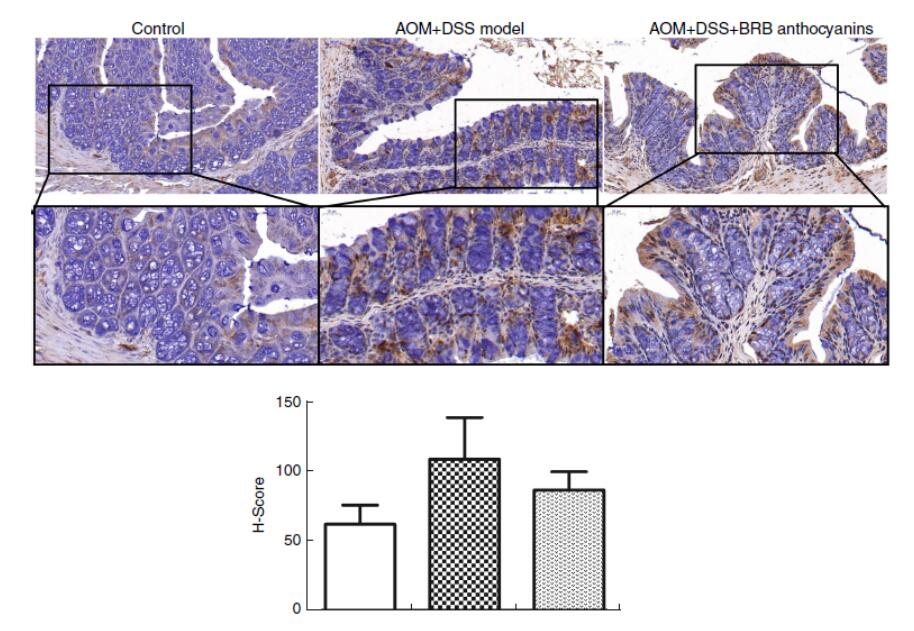 图3 黑树莓花色苷补充剂对小鼠结肠组织β-连环蛋白表达的影响。用免疫组化染色法(抗β-连环蛋白抗体1:100稀释)检测偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导小鼠的肠上皮细胞中β-连环蛋白的原位表达(分别放大20倍和40倍)。
图3 黑树莓花色苷补充剂对小鼠结肠组织β-连环蛋白表达的影响。用免疫组化染色法(抗β-连环蛋白抗体1:100稀释)检测偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导小鼠的肠上皮细胞中β-连环蛋白的原位表达(分别放大20倍和40倍)。 ,对照组;
,对照组; ,AOM+DSS模型;
,AOM+DSS模型; ,AOM+DSS+BRB花色苷。
,AOM+DSS+BRB花色苷。
恢复miR-24-1-5p表达对大肠癌细胞增殖和迁移的影响
为了进一步评价miR-24-1-5p的生物学功能,用miR-24-1-5p转染人HCT-116和Caco-2细胞,MTT法测定细胞的活力。与相应的对照细胞(图4(a)和(b))相比,miR-24-1-5p-转染的HCT-116和Caco-2细胞均表现出明显的细胞活力降低(HCT-116为56%,Caco-2为52%)。这些细胞也表现出显著的硬化减少,如伤口愈合试验所示(图4(c)和(d))。此外,它们表现出显著的细胞迁移和细胞存活率降低,分别由Transwell分析(图4(e)和(f))和菌落形成分析(图4(g)和(h))显示。综上所述,结果表明,过表达miR-24-1-5p可显著降低这些人结直肠癌细胞株的增殖、迁移和存活,从而表明miR-24-1-5p可能通过减少或抑制肿瘤细胞的生长而在结直肠癌中起到肿瘤抑制作用。
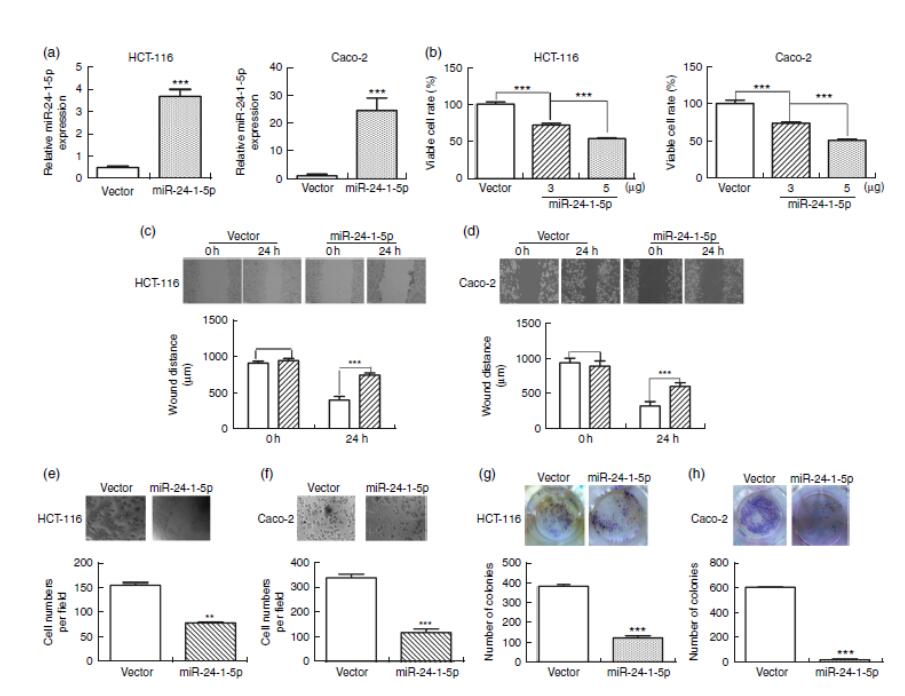
图4 miR-24-1-5p抑制人结直肠癌细胞的增殖、迁移和形成。用5μg的miR-24-1-5p转染HCT-116和Caco-2细胞24小时,然后进行以下分析:(a)用定量RT-PCR法测定miR-24-1-5p的表达水平。(b)用 3-(4,5-二甲基-2-噻唑)-2,5-二苯基四氮唑溴盐分析法测定细胞活力。(c,d)通过伤口愈合试验评估细胞迁移。对细胞迁移的抑制作用通过24小时伤口面积增加的百分比来确定,0小时伤口面积为100%。 ,空白载体;
,空白载体; ,miR-24-1-5p。(e,f)细胞侵袭用显微镜检查。(g,h)细胞群落形成的典型显微照片和定量分析。采用单因素方差分析比较组内和组间的差异,然后进行Fisher最小显著性差异分析。值是平均值(n=3),其标准误差由竖线表示。与空白载体对照组相比,平均值有显著性差异:**P<0.01,***P<0.001。
,miR-24-1-5p。(e,f)细胞侵袭用显微镜检查。(g,h)细胞群落形成的典型显微照片和定量分析。采用单因素方差分析比较组内和组间的差异,然后进行Fisher最小显著性差异分析。值是平均值(n=3),其标准误差由竖线表示。与空白载体对照组相比,平均值有显著性差异:**P<0.01,***P<0.001。
miR-24-1-5p调节β-连环蛋白
利用生物信息学方法预测了miR-24-1-5p可能结合的靶基因,采用Miranda 3.3a版默认参数和截止值(评分S≥140,能量E≤-7.0)。考虑到β-连环蛋白在CRC发展中的重要作用,选择β-连环蛋白作为可能的靶点。在β-连环蛋白的3′-UTR处检测到一个结合域,它可能与miR-24-1-5p相互作用(图5(a))。此外,为了研究miR-24-1-5p对β-连环蛋白表达的影响,过表达人HCT-116和Caco-2细胞中的miR-24-1-5p,用qRT-PCR和蛋白质印迹法检测β-连环蛋白水平的变化。在这两个细胞系中,当miR-24-1-5p过度表达时,β-连环蛋白的蛋白水平显著降低,而其mRNA水平保持不变(图5(b)和(c))。
进一步实验用miR-24-1-5p或对照载体转染HCT-116和Caco-2细胞24小时,在细胞进行蛋白质印迹分析前,将放线菌酮(10μM)在不同时间间隔加入细胞,测定β-连环蛋白的蛋白质合成率。结果表明,与对照质粒转染的细胞相比,miR-24-1-5p质粒转染细胞中β-连环蛋白的蛋白水平明显降低,提示miR-24-1-5p能促进β-连环蛋白的降解,影响其稳定性。结果还提示,miR-24-1-5p可能通过直接结合β-连环蛋白的3′-UTR来负向调节β-连环蛋白。
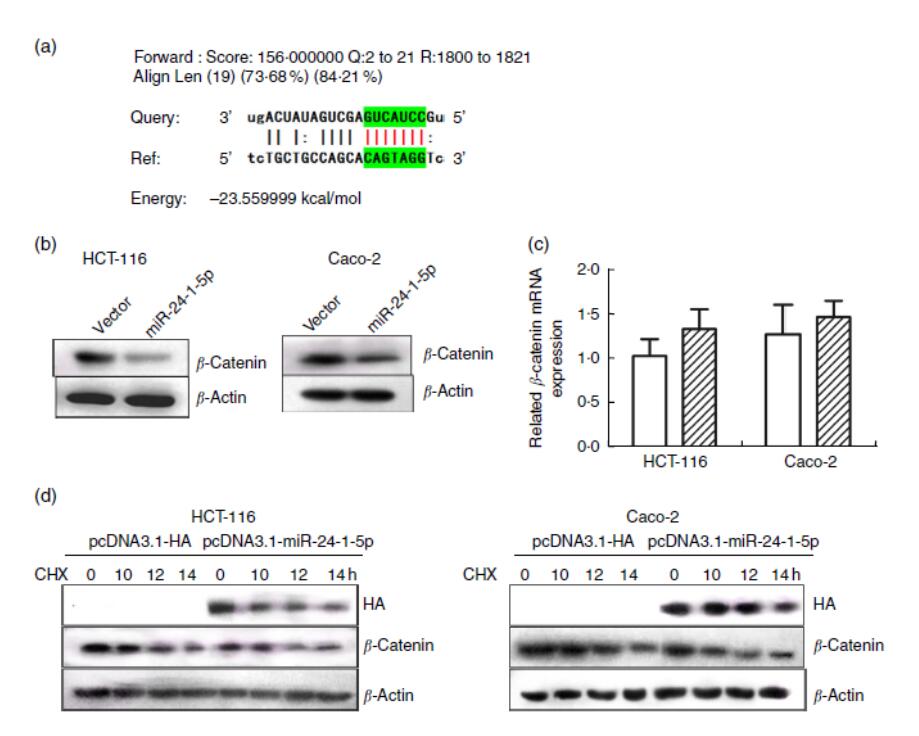 图5 miR-24-1-5p通过直接结合3′-UTR(未翻译区域)抑制β-连环蛋白的表达。(a)β-连环蛋白3′-UTR中假定的miR-24-1-5p结合位点的序列比对。(b)HCT-116和Caco-2细胞经miR-24-1-5p转染后β-连环蛋白水平的蛋白质印迹分析。(c)HCT-116和Caco-2细胞经miR-24-1-5p转染后β-连环蛋白水平的定量RT-PCR分析。(d)10μM放线菌酮(CHX)不同时间处理后β-连环蛋白水平(0、10、12、14小时)。采用单因素方差分析比较组内和组间的差异,然后进行Fisher最小显著性差异分析。
图5 miR-24-1-5p通过直接结合3′-UTR(未翻译区域)抑制β-连环蛋白的表达。(a)β-连环蛋白3′-UTR中假定的miR-24-1-5p结合位点的序列比对。(b)HCT-116和Caco-2细胞经miR-24-1-5p转染后β-连环蛋白水平的蛋白质印迹分析。(c)HCT-116和Caco-2细胞经miR-24-1-5p转染后β-连环蛋白水平的定量RT-PCR分析。(d)10μM放线菌酮(CHX)不同时间处理后β-连环蛋白水平(0、10、12、14小时)。采用单因素方差分析比较组内和组间的差异,然后进行Fisher最小显著性差异分析。 ,空白载体;
,空白载体; ,miR-24-1-5p。HA,血凝素。
,miR-24-1-5p。HA,血凝素。
与Wnt/β-连环蛋白通路相关的基因由miR-24-1-5p调控
还研究了miR-24-1-5p过度表达对β-连环蛋白信号通路下游几个靶基因的影响。用蛋白质印迹和qRT-PCR检测HCT-116和Caco-2细胞周期蛋白D1、c-Myc和CDK4表达水平的变化。HCT-116和Caco-2细胞中这些基因的mRNA和蛋白质水平在被miR-24-1-5p转染后24小时均下降(图6(a)和(b))。此外,还分析了与Wnt/β-连环蛋白通路相关的其他基因,如SFRP2、SFRP5、E-钙粘蛋白、GSK3-β、P-GSK3-β和Bcl-2。E-钙粘蛋白、p-GSK3-β和SFRP5的表达上调,Bcl-2的表达下调,GSK3-β和SFRP2的表达无显著变化(图6(c))。总之,来自体外人类癌细胞的数据表明β-连环蛋白可能是miR-24-1-5p的直接靶点,miR-24-1-5p的调节作用通过影响β-连环蛋白下游靶基因表达来发挥。
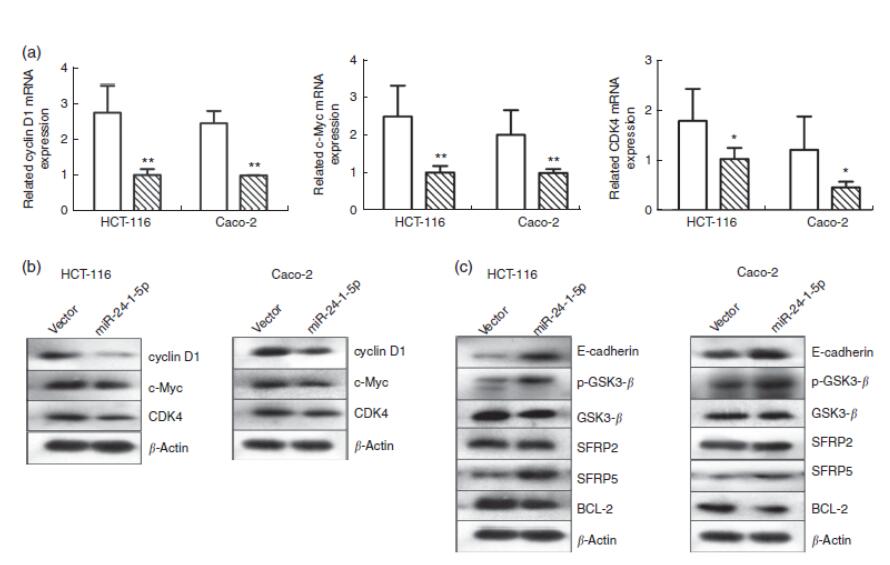
图6 miR-24-1-5p对结直肠癌细胞Wnt/β-连环蛋白信号相关基因的影响。HCT-116和Caco-2细胞在5μg miR-24-1-5p转染24h后,细胞周期蛋白D1、c-Myc 和细胞周期蛋白依赖性激酶4(CDK4)的mRNA定量RT-PCR(a)或蛋白质印迹(b)分析结果。(c)经miR-24-1-5p转染24小时后HCT-116和Caco-2细胞中糖原合成酶激酶3β(GSK3-β)、分泌型卷曲相关蛋白2(SFRP2)、分泌型卷曲相关蛋白5(SFRP5)、B细胞淋巴瘤2(Bcl-2)、E-钙粘蛋白和磷酸糖原合成酶激酶3β(p-GSK3-β)蛋白水平的蛋白质印迹分析。组内和组间差异采用单因素方差分析比较,然后进行Fisher最小显著性分析。与空白载体对照组相比,平均值有显著差异:*P<0.05,**P<0.01。 ,空白载体;
,空白载体; ,miR-24-1-5p。
,miR-24-1-5p。
讨论
积累的证据表明,细胞中严格调控的RNA网络可能被异常表达的miRNA破坏,从而引发癌症的发展和转移。miRNA在各种人类癌症中的重要性表明,调节miRNA的表达可能是癌症化学预防和治疗的新策略(36)。在本研究中,食用含有BRB花色苷饮食的AOM/DSS诱导小鼠中,miR-24-1-5p显著上调。此外,机制研究表明,miR-24-1-5p可能起到肿瘤抑制作用,该作用由其在人类CRC细胞中过表达时抑制细胞增殖、迁移、存活以及群落形成的能力产生(图3)。miR-24-1-5p的肿瘤抑制作用似乎是由对β-连环蛋白信号通路的影响所介导(图4和5)。
Wnt/β-连环蛋白信号通路在癌症中的作用已通过该信号通路对CRC细胞生长、代谢、血管生成、迁移和转移的影响得到证实(37)。β-连环蛋白及其下游靶基因(包括c-Myc、细胞周期蛋白D1和CDK4)的异常激活可加速CRC的发生和发展。我们的结果清楚地表明,在人类结直肠癌细胞中,这些基因可以通过改变miR-24-1-5p的表达来调节。实际上,在HCT-116和Caco-2细胞中miR-24-1-5p的过表达下调了β-连环蛋白、c-Myc、细胞周期蛋白D1和CDK4的表达(图6)。阐明经β-连环蛋白介导的miR-24-1-5p调控的肿瘤通路和靶点,将为人类结直肠癌的发生过程提供新的认识。不幸的是,由于伦理问题,我们无法从人类受试者那里获得直接证据来支持我们的主张。然而,有必要进一步研究确定在CRC中靶向miR-24-1-5p的可行性,将其作为临床水平对抗CRC的一种经济有效且简单的治疗策略。BRB花色苷作为一种CRC的化学预防剂已被多次研究。该方向在动物模型和临床病人中得到了广泛研究。BRB花色苷的化学预防作用机制包括诱导细胞凋亡和分化,减少细胞增殖、炎症、血管生成和侵袭性(38)。BRB花色苷对结直肠癌的化学预防作用似乎也涉及肠道微生物群的改变和DNA去甲基化(39)。在本研究中,我们首次证明了BRB花色苷对CRC的化学预防作用也可能涉及到对miR-24-1-5p的上调,且我们通过小鼠模型证明了这一点。
总之,miR-24-1-5p可能作为肿瘤抑制因子,通过下调β-连环蛋白及其下游靶基因的表达来调控CRC细胞的生长。这可能是BRB花色苷显示其对CRC的化学预防作用的机制的一部分。因此,miR-24-1-5p可被认为是未来预防或治疗结直肠癌的潜在药物。
作为重要的表观遗传调控因子,microRNA通过触发靶mRNA的降解和/或通过抑制其翻译来调节蛋白质的表达。已报道在包括结直肠癌的一些癌症中存在microRNA表达失调。在这项研究中,microRNA阵列差异分析显示,在用黑树莓花色苷喂养9周的偶氮甲烷/葡聚糖硫酸钠诱导的小鼠结肠组织中,miR-24-1-5p的表达显著增强。在人类结直肠癌细胞中,miR-24-1-5p的过度表达显著抑制了β-连环蛋白的表达,同时降低了细胞的增殖、迁移和存活。此外,mir-24-1-5p可以靶向β-连环蛋白并触发β-连环蛋白及其下游靶基因的负调控环。β-连环蛋白信号对人结直肠癌的形成和发展至关重要。因此,目前的研究发现,miR-24-1-5p是β-连环蛋白的有效调节因子,这可能为β-连环蛋白信号驱动的结直肠癌提供一种新的化学预防和治疗策略。
结直肠癌(CRC)是全球第三大恶性肿瘤,约占所有癌症发病率和死亡率的10 %。尽管过去20年里人们致力于CRC诊断和治疗策略,但仍有五分之一的患者被诊断为CRC时已进入晚期,通常采用手术和辅助化疗相结合的方法进行治疗(3-5)。大多数患者发生化疗耐药,癌症可能再次发生在这些患者身上并最终转移(6)。最近一些研究已经报道了microRNA(miRNA)表达与CRC(7-9)进展的显著关联。因此,在RNA网络分析的基础上,寻找一种可用于预防或治疗结直肠癌的新靶点是十分必要的。
miRNA是一种长度为18-22 nt的内源性非编码小RNA,通过结合mRNA的3′-UTR(未翻译区域)来干扰其翻译,从而调节基因表达,直接降解结合的mRNA(10,11)。在许多人类癌症中,miRNA的异常表达可激活肿瘤抑制基因或癌基因。在结直肠癌中,有几个miRNA被鉴定为肿瘤发育的必要条件。Michael等人(12)首次报道了人结直肠癌组织和正常结肠直肠粘膜中miRNA的表达差异,共检测到28个差异表达的miRNA,包括miR-320、miR-321、miR-200c、miR-223和miR-145。Monzo等人(13)发现,miR-17-5p可以靶向E2F转录因子1,该转录因子存在于人类结肠早期胚胎发育和肿瘤发生时期并调节细胞增殖。其他研究者报道,miR-34a直接并负向控制p53的一系列下游靶基因,形成miR-34a-p53的正反馈环(14)。Bandres等人(15)发现,miRNA-451可以调节迁移抑制因子(MIF)的表达,从而抑制CRC细胞的增殖,提高肿瘤对放疗的敏感性。此外,miRNA-320a和miR-139可分别靶向β-连环蛋白和RAP1B(RAS癌基因家族成员),抑制人结肠癌细胞的增殖(16,17)。2001年,Lagos Quintana等人(18)在研究非脊柱动物和脊柱动物时首次发现了miR-24。人的miR-24由miR-24-1和miR-24-2组成,分别位于9号和19号染色体上,成熟序列包括miR-24-3p、miR-24-1-5p和miR-24-2-5p(19)。有报道称,miR-24-1在数种癌症组织中表达异常(20-26)。然而,miR-24-1-5p在CRC中的功能尚不清楚。
黑树莓(BRB)属于莓类中的Rubus occidentalis科,富含黄酮类和酚酸(27,28)。BRB对结直肠癌的化学预防作用已被本研究组和其他研究组(29-32)的研究所证实。然而,很少有研究关注miRNA与BRB在结直肠癌中的化学预防作用及其潜在机制的相关性。
在本研究中,我们假设miR-24-1-5p在结直肠癌的发生和发展中起重要作用,也参与了BRB对结直肠癌的化学预防作用。为了验证这一假设,我们在小鼠上进行了一项体内研究,并对人类细胞系进行了数项体外研究,发现miR-24-1-5p可以作为肿瘤抑制因子发挥作用,其靶向数个CRC相关通路中的β-连环蛋白相关癌基因。我们目前的发现可能有助于发展新的诊断和治疗策略,用以预防和治疗结直肠癌。
方法
化合物与试剂
氧化偶氮甲烷(AOM,A5486)购自Sigma Aldrich公司,葡聚糖硫酸钠(DSS,0216011091)购自MP公司。建丰自然产品技术有限公司提供BRB花色苷(纯度>90%)。BRB花色苷由三种主要花色苷——矢车菊素葡萄糖苷、矢车菊素木糖基芦丁糖苷和矢车菊素芸香糖苷组成,含量分别为2.63、0.73和16.91 mg/g。矢车菊素桑布双糖苷的含量不太丰富,我们最近发表的论文(33)详细介绍了这一点。β-连环蛋白、细胞周期蛋白D1、c-Myc和细胞周期蛋白依赖性激酶4(CDK4)从细胞信号技术公司获得,β-肌动蛋白、E-钙粘蛋白、糖原合酶激酶3β(GSK3-β)、磷酸糖原合酶激酶3β(p-GSK3-β)、B细胞淋巴瘤-2(BCL-2)和血凝素从Sangon Biotech有限公司获得。分泌型卷曲相关蛋白2(SFRP2)购自Abcam公司。从Biorbyt Explore生物试剂公司中购得分泌型卷曲相关蛋白5(SFRP5)。免疫印迹的二级抗体为辣根过氧化物酶偶联的山羊抗兔和山羊抗鼠IgG,从ComWin生物科技有限公司获得。
动物护理和实验方案
5周大的雄性C57BL/6J小鼠(18-20g,本溪长生实验动物技术有限公司)给予随意饮水,维持在12小时光暗循环下。这些动物被关在笼子里(每笼5只动物),房间温度设置为21±2.0°C,湿度控制50±5%。所有动物实验方案均获辽宁中医药大学伦理委员会(中国,沈阳)批准。
结肠炎诱导的结直肠癌小鼠模型的建立及黑树莓花色苷的补充
在1周适应期后,小鼠单次腹腔注射AOM(10 mg/kg体重)。1周后,在饮用水中给予2% DSS持续1周,接着给予正常水持续2周,该循环再重复2次。
小鼠分为三组:一组健康对照组和两组AOM/DSS处理组。每组10只。健康对照组的小鼠没有接受任何处理,喂以饲料超过9周。对于两个AOM/DSS处理组,一组只用饲料喂养,而另一组则用含有7.0μmol/g BRB花色苷的饲料(10%BRB)喂养。添加饲料中BRB花色苷浓度相当于10%冻干BRB粉中的花色苷含量,这是基于前人研究(32,34)选定的。饲料和BRB花色苷补充饲料的配方组成如表1所示。实验中的所有饲料使用前储存于-20 ℃。
在第9周结束后,小鼠吸入二氧化碳后颈椎脱臼处死。结肠组织立即冷冻在液氮中,然后储存在-80℃下准备进行microRNA阵列分析。
表1 饲料和BRB花色苷补充饲料的配方
MicroRNA微阵列表达谱与数据分析
miRNA表达谱由Agilent Mouse miRNA(8×60K,设计ID:070155;Agilent Technologies公司)生成。总RNA用NanoDrop ND-2000(Thermo Scientific)定量。用Agilent Bioanalyzer 2100(Agilent Technologies公司)评估RNA完整性。总RNA去磷酸化、变性后,用花青苷-3-胞苷三磷酸标记。纯化后,标记的RNA与微阵列杂交。清洗后,使用Agilent Scanner G2505C(Agilent Technologies公司)对阵列进行扫描。阵列图像用Feature Extraction软件(版本10.7.1.1;Agilent Technologies公司)获取。使用GeneSpring软件(版本13.1;Agilent Technologies公司)分析原始数据。
RNA提取与定量实时PCR
总RNA使用Trizol试剂盒(Comwin Biotech有限公司)从细胞中分离,并使用miRNA cDNA合成试剂盒(ComWin Biotech有限公司)通过逆转录产生第一链互补DNA(cDNA)编码的带poly A尾的miRNA。实时PCR(RT-PCR)采用SYBR Green方法以特异性引物进行。根据制造商的说明,RT-PCR在Applied Biosystems 7500实时PCR系统上进行扩增,并用microRNA实时分析试剂盒(ComWin Biotech有限公司)进行分析。引物序列为:mmu-miR-24-1-5p:正链5′-AGTGCCTACTGAGCTGATATCAGT-3′;hsa-miR-24-1-5p:正链5′-GTGCCTACTGAGCTGATATCAGT-3′;β-连环蛋白:正链 5′-GCTGGTGACAGGGAAGACAT-3′,反链5′-AGGCAAACCGTGAAAAGATG-3′;细胞周期蛋白D1:正链5′-AACTACCTGGACCGCTTCCT-3′,反链 5′-CCACTTGAGCTTGTTCACCA-3′;c-Myc:正链5′-TTCGGGTAGTGGAAAACCAG-3′,反链5′-CAGCAGCTCGAATTTCTTCC-3′;CDK4:正链 5′-ATTGGTGTCGGTGCCTATG-3′,反链5′-AACTGTGCTGATGGGAAGG-3′。这些基因的表达水平按照2–ΔΔCT方法以内部对照(U6:正链 5′-ATTGGAACGATACAGAGAAGATT-3′,反链5′-GGAACGCTTCACGAATTTG-3′或β-肌动蛋白:正链5′-AGGCAAACCGTGAAAAGATG-3′,反链 5′-AGGCAAACCGTGAAAAGATG-3′)正常化。
(译者注:正链(forward strand),与mRNA序列相同的DNA单链,另一条则为反链(reverse strand);互补DNA链中携带编码蛋白质信息的链称为正义链(sense strand),又称编码链(coding strand),另一条称为反义链(antisense strand),又称模板链(template strand)。)
质粒构建和转染
miR-24-1-5p的成熟序列为UGCCUACUGAGCUGAUAUCAGU。人pri miR-24-1-5p序列构建使用如下引物:miR-24-1-5p正链, 5′-CCGCTCGAGCAACAGGGTTTTCCAAGTCTAC-3′;miR-24-1-5p反链,5′-GGAAGATCTTCACCTAAGTCGGAAATCATGTGGTA-3′。PCR产物用XhoI和BglII 这一对内切酶消化,插入到XhoI-BglII切开的pcDNA3-血凝素(质粒载体)中。然后用lipofectamine 2000(Invitrogen)将HCT-116和Caco-2细胞与人pri-miR-24-1-5p一起转染。用空白质粒转染的细胞作为阴性对照。构建的人pri-miR-24-1-5p经测序验证。
(备注:成熟的miRNAs是由较长的初级转录物经过一系列核酸酶的剪切加工而产生的,初级转录物称为pri-miRNA。pri-miRNA长度从几百到几千个碱基不等,带有5‘帽子和3’poly A尾巴,以及1到数个发夹径环结构。pri-miRNA经剪切产生约70个碱基的miRNA前体,即pre-miRNA。pre-miRNA为单一发夹结构,5‘带有磷酸基团,3’有两个突出碱基,并带有3‘羟基。pre-miRNA经进一步剪切,形成长度约为22个碱基的单链成熟miRNA。)
细胞培养
人结直肠癌细胞系HCT-116(CCL-247;美国模式培养物集存库(ATCC,American type culture collection))和Caco-2(北京癌症研究所细胞库)分别在Dulbecco改进的Eagle's培养基和α-最低基础培养基(Hyclone,Thermo Fisher Scientific公司)中培养。这两种培养基中都添加10%(v/v)胎牛血清(大连美伦生物)、10000 U/ml青霉素G、10 mg/ml链霉素,在37℃的5% CO2湿润空气中孵育。
伤口愈合试验
伤口愈合试验采用汇合度为70%的HCT-116和Caco-2细胞。首先,用无菌移液管尖端在细胞单层上划一条直线。用PBS洗涤细胞,然后加入新鲜培养基。然后用miR-24-1-5p转染,24小时后,丢弃培养基,用显微照片计算划痕中剩余的间隙。
细胞增殖和群落形成分析
用 3-(4,5-二甲基-2-噻唑)-2,5-二苯基四氮唑溴盐(MTT)法测定细胞活力。简单来说,在miR-24-1-5p转染24小时后,用0.5 mg/ml MTT试剂培养Caco-2和HCT-116细胞4小时。然后去除培养基,在每个孔中加入100μl二甲亚砜,以溶解甲赞晶体。在490nm处测量板的吸光度。为了进一步研究miR-24-1-5p在癌细胞存活中的作用,将细胞以200/孔接种在六孔板中,培养2周进行菌落形成分析。出现的细胞群落首先用PBS洗涤,然后用甲醇固定,再用0.5%的吉姆萨染液(Giemsa,天青-伊红)染色。用倒置显微镜计数每孔中出现的群落数。
Transwell迁移分析
将Caco-2和HCT-116细胞接种在Transwell孔的上室,在下室中加入无血清培养基。24小时后,用棉签去除膜顶部的细胞,然后用4%多聚甲醛固定Transwell孔插入膜。迁移到膜底的细胞用4’,6-二脒基-2-苯基吲哚(北京鼎国昌盛生物技术有限公司)染色。从每个插入膜中随机选择10个域,在20倍放大的显微镜下进行捕获。使用Image Pro Plus 6.0对细胞数进行量化。为了计算迁移指数,将每个处理组捕获的细胞数归一化为对照组细胞数。
microRNA的靶向预测
利用Miranda 3.3a版的算法和网站工具(http://www.microrna.org/microrna/home.do)对miR-24-1-5p的靶基因进行了预测,并给出了默认参数和截止值(评分S≥140,能量E≤-7.0)。
蛋白质印迹
细胞在含1 mmol/L苯甲磺酰氟(北京鼎国昌盛生物技术有限公司)的冷冻放射免疫沉淀分析缓冲液中置于冰上30分钟。在4℃下14000 g离心15 分钟,收集上清液,用10%凝胶SDS-PAGE法测定含有40μg蛋白的上清液。电泳后,将凝胶中的蛋白质转移到硝酸纤维素膜((Sangon Biotech有限公司)上,用5%牛血清白蛋白V(Roche)在Tris缓冲盐水和吐温20缓冲液中封闭2小时,然后在室温下与合适的一级抗体孵育3小时,然后上相应的二级抗体。用增强化学发光技术(Amersham Life Science)检测印迹上的信号。
免疫组织化学染色
收集AOM/DSS诱导小鼠肠道组织,无论是否含有BRB花色苷,固定在10%的福尔马林缓冲液中,并如前所述嵌入石蜡中(35)。组织切片(4μm厚)用抗E-钙粘蛋白抗体(Santa Cruz生物技术)进行免疫组化染色。流程与先前报道类似(35)。简言之,切片脱蜡、复水,3% H2O2淬灭,10%的正常血清封闭,然后用β-连环蛋白抗体在4℃孵育过夜,接着用二级抗体和亲和素-生物素复合物(ABC试剂盒;Vector Laboratory)孵育。用3’,5’-二氨基联苯胺进行染色,再用苏木精进行轻微复染。彩色载玻片的图像由Aperio Image系统采集。
统计方法
采用SPSS 19.0统计软件对数据进行分析。为了比较组内和组间的差异,采用t检验和方差分析,然后进行Fisher最小显著性差异分析。在P<0.05水平上考虑统计学意义。
结论
组织microRNA微阵列分析
为了阐明miRNA介导的结直肠癌肿瘤发生通路,识别异常表达的miRNA是第一步。基于这一点,我们进行了组织miRNA微阵列分析,以获得AOM/DSS诱导小鼠(无论是否补充BRB花色苷)结肠组织中总miRNA表达特征。两组结果的比较表明,通过给予BRB花色苷,数个miRNA的调节有差异(图1(a))。组织阵列的层次聚类分析结果如图1(b)所示。AOM/DSS诱导组表达低水平的miR-24-1-5p,但用BRB花色苷喂养9周后,miR-24-1-5p水平显著上调,提示miR-24-1-5p可能对肿瘤形成有抑制作用。这也证实了BRB花色苷的化学预防功能。
miR-24-1-5p在偶氮甲烷/葡聚糖硫酸钠诱导的小鼠结肠组织中表达下调,在用黑树莓花色苷处理的人结直肠癌细胞系中表达上调
为了进一步评估小鼠模型微阵列分析结果,采用定量RT-PCR(qRT-PCR)分析了有与无BRB花色苷给药的AOM/DSS诱导组结肠组织中的miR-24-1-5p表达。qRT-PCR获得的miR-24-1-5p的表达模式与微阵列数据一致(图2(a))。在用不同剂量的BRB花色苷处理后,测定了几种人结直肠癌细胞系(Caco-2、HCT-116、Lovo、HT29和SW480)中miR-24-1-5p的表达。BRB花色苷导致miR-24-1-5p水平升高。在BRB花色苷处理的人类肿瘤细胞中,miR-24-1-5p的表达模式与用BRB花色苷补充饮食喂养的小鼠结肠组织获得的miRNA微阵列和qRT-PCR数据一致(图2(b))。
黑树莓花色苷可降低偶氮甲烷/葡聚糖硫酸钠诱导小鼠的β-连环蛋白含量。
为了研究β-连环蛋白在BRB花色苷原位调节的AOM/DSS诱导小鼠中的作用,我们进行了免疫组化染色。与不给予BRB花色苷饮食的AOM/DSS诱导小鼠相比,给予BRB花色苷饮食的AOM/DSS诱导小鼠小肠的β-连环蛋白水平显著降低(图3)。
恢复miR-24-1-5p表达对大肠癌细胞增殖和迁移的影响
为了进一步评价miR-24-1-5p的生物学功能,用miR-24-1-5p转染人HCT-116和Caco-2细胞,MTT法测定细胞的活力。与相应的对照细胞(图4(a)和(b))相比,miR-24-1-5p-转染的HCT-116和Caco-2细胞均表现出明显的细胞活力降低(HCT-116为56%,Caco-2为52%)。这些细胞也表现出显著的硬化减少,如伤口愈合试验所示(图4(c)和(d))。此外,它们表现出显著的细胞迁移和细胞存活率降低,分别由Transwell分析(图4(e)和(f))和菌落形成分析(图4(g)和(h))显示。综上所述,结果表明,过表达miR-24-1-5p可显著降低这些人结直肠癌细胞株的增殖、迁移和存活,从而表明miR-24-1-5p可能通过减少或抑制肿瘤细胞的生长而在结直肠癌中起到肿瘤抑制作用。
miR-24-1-5p调节β-连环蛋白
利用生物信息学方法预测了miR-24-1-5p可能结合的靶基因,采用Miranda 3.3a版默认参数和截止值(评分S≥140,能量E≤-7.0)。考虑到β-连环蛋白在CRC发展中的重要作用,选择β-连环蛋白作为可能的靶点。在β-连环蛋白的3′-UTR处检测到一个结合域,它可能与miR-24-1-5p相互作用(图5(a))。此外,为了研究miR-24-1-5p对β-连环蛋白表达的影响,过表达人HCT-116和Caco-2细胞中的miR-24-1-5p,用qRT-PCR和蛋白质印迹法检测β-连环蛋白水平的变化。在这两个细胞系中,当miR-24-1-5p过度表达时,β-连环蛋白的蛋白水平显著降低,而其mRNA水平保持不变(图5(b)和(c))。
进一步实验用miR-24-1-5p或对照载体转染HCT-116和Caco-2细胞24小时,在细胞进行蛋白质印迹分析前,将放线菌酮(10μM)在不同时间间隔加入细胞,测定β-连环蛋白的蛋白质合成率。结果表明,与对照质粒转染的细胞相比,miR-24-1-5p质粒转染细胞中β-连环蛋白的蛋白水平明显降低,提示miR-24-1-5p能促进β-连环蛋白的降解,影响其稳定性。结果还提示,miR-24-1-5p可能通过直接结合β-连环蛋白的3′-UTR来负向调节β-连环蛋白。
与Wnt/β-连环蛋白通路相关的基因由miR-24-1-5p调控
还研究了miR-24-1-5p过度表达对β-连环蛋白信号通路下游几个靶基因的影响。用蛋白质印迹和qRT-PCR检测HCT-116和Caco-2细胞周期蛋白D1、c-Myc和CDK4表达水平的变化。HCT-116和Caco-2细胞中这些基因的mRNA和蛋白质水平在被miR-24-1-5p转染后24小时均下降(图6(a)和(b))。此外,还分析了与Wnt/β-连环蛋白通路相关的其他基因,如SFRP2、SFRP5、E-钙粘蛋白、GSK3-β、P-GSK3-β和Bcl-2。E-钙粘蛋白、p-GSK3-β和SFRP5的表达上调,Bcl-2的表达下调,GSK3-β和SFRP2的表达无显著变化(图6(c))。总之,来自体外人类癌细胞的数据表明β-连环蛋白可能是miR-24-1-5p的直接靶点,miR-24-1-5p的调节作用通过影响β-连环蛋白下游靶基因表达来发挥。
讨论
积累的证据表明,细胞中严格调控的RNA网络可能被异常表达的miRNA破坏,从而引发癌症的发展和转移。miRNA在各种人类癌症中的重要性表明,调节miRNA的表达可能是癌症化学预防和治疗的新策略(36)。在本研究中,食用含有BRB花色苷饮食的AOM/DSS诱导小鼠中,miR-24-1-5p显著上调。此外,机制研究表明,miR-24-1-5p可能起到肿瘤抑制作用,该作用由其在人类CRC细胞中过表达时抑制细胞增殖、迁移、存活以及群落形成的能力产生(图3)。miR-24-1-5p的肿瘤抑制作用似乎是由对β-连环蛋白信号通路的影响所介导(图4和5)。
Wnt/β-连环蛋白信号通路在癌症中的作用已通过该信号通路对CRC细胞生长、代谢、血管生成、迁移和转移的影响得到证实(37)。β-连环蛋白及其下游靶基因(包括c-Myc、细胞周期蛋白D1和CDK4)的异常激活可加速CRC的发生和发展。我们的结果清楚地表明,在人类结直肠癌细胞中,这些基因可以通过改变miR-24-1-5p的表达来调节。实际上,在HCT-116和Caco-2细胞中miR-24-1-5p的过表达下调了β-连环蛋白、c-Myc、细胞周期蛋白D1和CDK4的表达(图6)。阐明经β-连环蛋白介导的miR-24-1-5p调控的肿瘤通路和靶点,将为人类结直肠癌的发生过程提供新的认识。不幸的是,由于伦理问题,我们无法从人类受试者那里获得直接证据来支持我们的主张。然而,有必要进一步研究确定在CRC中靶向miR-24-1-5p的可行性,将其作为临床水平对抗CRC的一种经济有效且简单的治疗策略。BRB花色苷作为一种CRC的化学预防剂已被多次研究。该方向在动物模型和临床病人中得到了广泛研究。BRB花色苷的化学预防作用机制包括诱导细胞凋亡和分化,减少细胞增殖、炎症、血管生成和侵袭性(38)。BRB花色苷对结直肠癌的化学预防作用似乎也涉及肠道微生物群的改变和DNA去甲基化(39)。在本研究中,我们首次证明了BRB花色苷对CRC的化学预防作用也可能涉及到对miR-24-1-5p的上调,且我们通过小鼠模型证明了这一点。
总之,miR-24-1-5p可能作为肿瘤抑制因子,通过下调β-连环蛋白及其下游靶基因的表达来调控CRC细胞的生长。这可能是BRB花色苷显示其对CRC的化学预防作用的机制的一部分。因此,miR-24-1-5p可被认为是未来预防或治疗结直肠癌的潜在药物。
本文由福山生物整理翻译,转载请注明出处。