






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
黑树莓花色苷化学预防结直肠癌的研究——肠道微生物的调节和SFRP2去甲基化
发表于:2019-02-15 作者:admin 来源:本站 点击量:16567

摘要:
冻干黑树莓粉(BRB)被认为是一种潜在的癌症化学预防制剂。在本研究中,我们对AOM/DSS处理C57BL/6J小鼠进行了为期12周含BRB花色苷的饮食干预,最终降低了结肠癌的发生。与没有接受BRB花色苷干预的AOM / DSS处理小鼠相比,这些动物一直具有较低的肿瘤多样性。在AOM / DSS处理小鼠中,包括脱硫弧菌(Desulfovibrio sp)和肠球菌(Enterococcus spp)在内的病原菌的数量显著增加,而直肠真杆菌(Eubacterium rectale)、柔嫩梭菌群(Faecalibacterium prausnitzii)、乳酸菌(Lactobacillus)等益生菌显著降低,但补充BRB花色苷可以逆转肠道微生物的这种不平衡。BRB花色苷也引起SFRP2基因启动子的去甲基化,使mRNA和蛋白水平SFRP2的表达增加。此外,在这些动物中,BRB花色苷下调DNMT31、DNMT3B以及p-STAT3的表达水平。总之,这些结果表明,BRB花色苷可以调节肠共生菌群的组成和炎症的变化,而SFRP2基因的甲基化状态可能在结直肠癌的化学预防中发挥核心作用。
关键词:BRB花色苷;肠道菌群;甲基化改变;SFRP2;p-STAT3
1.背景介绍
人类的大肠是非常活跃的发酵部位,并且共生着不同种类的细菌,尤其是结肠中菌群数量最多(每克粪便多达1012个细菌)(1)。这些大部分细菌属于厚壁菌和拟杆菌属,参与许多生理活动和新陈代谢,包括食物的消化、为结肠上皮细胞供应能量和调节免疫应答(2)。这种细菌群落的组成不仅在个体间有很大的差异,而且它也是一个动态群落,容易受饮食因素和多种疾病条件的影响。肠道菌群的成分变化可以改变肠道的通透性,导致炎症性肠病,甚至结直肠癌(CRC)的逐步发展。肠道菌群参与CRC形成和发展的机制尚不清楚。一种被普遍接受的理论是肠道菌群在炎症性肠病的发展中起着基础性作用,长期慢性炎症可显著增加CRC的风险(3)。因此,肠道菌群的改善被认为是预防炎症性肠病和CRC的潜在疗法。
CpG岛的DNA甲基化是一种主要的表观遗传修饰,与癌变密切相关(4)。在肿瘤发生过程中,一系列的基因被证明是超或低甲基化的(5)。先前已经观察到,结直肠癌中编码分泌的卷曲相关蛋白(SFRPs)的基因出现频繁的启动子超甲基化和基因沉默(6)。SFRPs具有与包含一组WNT拮抗剂的WNT受体卷曲蛋白中的结构域相似的结构域,因此它可以抑制WNT受体与其配体的结合并在发育过程中下调Wnt /β-catenin信号通路(7,8)。SPRF2是SFRP基因家族的成员。在CRC组织和某些结肠癌细胞系中,SFRP基因启动子的超甲基化可导致SFRPs的水平下降(如SFRP4和SFRP5)以及随后的异常WNT信号传导(9)。此外,DNA甲基化可能会受到慢性炎症的影响。例如,在幽门螺杆菌诱导的胃癌发生过程中,促炎细胞因子白细胞介素-1β(IL-1β)通过产生一氧化氮增强DNMT活性,导致CpG甲基化介导的基因沉默(10)。最常被接受的理论是肠道微生物、免疫系统和表观遗传修饰之间可能存在复杂而动态的关系(11)。
花色苷是植物化学物质,丰富地存在于各种各样的食物中,包括茶、咖啡、巧克力、葡萄酒、水果或深色的蔬菜(2)。食用丰富的蔬菜和水果可以降低各种癌症,特别是结肠癌的风险。以往的研究表明,膳食花色苷及其代谢物可以促进有益菌生长,同时抑制病原菌的生长,有助于维持肠道健康(12)。
BRB属于浆果的Rubus occidentalis家族,原产于美国。BRB的生物活性物质根据其化学结构分为酚酸、类黄酮、原花青素、鞣酸、苯乙烯、木脂素、萜烯和类固醇。BRB中活性最高的活性成分是鞣花酸和花色苷,能够克服许多化学致癌物质的作用,有效清除自由基。BRB冻干粉被认为是消化道癌症的一种可能的癌症化学预防剂(13)。然而,迄今为止,很少有关于BRB花色苷作用和作用机制的实验研究发表。
在本研究中,我们通过检测某些参与了致癌作用的肠道菌群有关成分变化和某些基因的表观遗传情况来研究BRB花色苷饮食对氧化偶氮甲烷(AOM)/葡聚糖硫酸钠(DSS)诱导的CRC小鼠模型的癌症化学预防作用。
2. 材料和方法
2.1 细胞系和试剂
于2011年获得人结肠癌细胞系HCT116(CCL-247,美国标准菌库,ATCC)和LoVo(北京癌症研究所细胞库,中国北京),保存为冷冻原种。所有的细胞系都在Dulbecco改良的Eagle's培养基(Hyclone,Thermo Fisher Scientific Inc.)中培养,并通过它们的核型和形态进行鉴定。培养基补充有10%胎牛血清和抗生素(10000U/ml青霉素,10μg/ml链霉素)。在37℃,含有5%的二氧化碳的培养箱中培养。
从BRB成熟果实中提取的BRB花色苷(纯度为> 90%),由中国天津建丰自然产品技术有限公司提供。 BRB花色苷由三大类花色苷——花色苷-O-葡糖苷、花色苷-O-木糖基葡糖苷和花色苷-O-芸香糖苷组成,其含量分别为2.63、0.73和16.91 mg/g,而花色苷-o-sambubioside含量则较不丰富(14)。AOM(批号SLBN5975V)购自美国Sigma-Aldrich公司。DSS(批号160110)购自美国MP公司。
2.2动物饲养和实验方案
将5周龄的C57BL / 6J小鼠(18-20g)维持在12h / 12h明暗周期并随意饮水。动物饲养在动物笼中,房间保持一定温度(21±2.0℃)和湿度(50%±5%)。所有动物实验方案经辽宁中医药大学伦理委员会(中国沈阳)批准。
2.3建立结肠炎诱导的CRC小鼠模型和BRB花色苷干预
AOM/DSS模型是公认的化学诱导小鼠结肠炎相关癌症模型。适应一周后,雄性小鼠(6周龄)接受单次腹腔内注射AOM(10mg/kg体重)。同时,每天给这些动物食用含有BRB花色苷的饮食。AOM注射1周后,给予动物1周含2%DSS的饮用水,然后给予正常饮用水2周,并重复该步骤两次(15)。将小鼠分成四组:一个对照组和三个AOM / DSS处理组。健康对照组(n = 10)C57 / BJ小鼠在12周期间给予普通饮食饲喂。三个AOM / DSS处理组小鼠(n = 11)每天喂食含或不含BRB花色苷的食物。饮食中BRB花色苷的浓度分别为3.5μmol/ g(LBA)和7.0μmol/ g(MBA),分别相当于5%和10%BRB冻干粉中花色苷的含量,这是在我们以往的研究基础上选择的浓度(13)。在实验中使用之前将饮食储存在-20℃。
2.4样品收集
在第12周结束时,收集动物全血并储存在-80℃。然后所有的动物吸入二氧化碳后颈椎脱臼处死。如前所述(16),通过在在37℃、15mM乙二胺四乙酸缓冲液中孵育解剖的肠组织30分钟来分离小鼠肠上皮细胞。从每只动物收集肿瘤组织、肠组织、上皮细胞和肠内容物,立即储存在-80℃,然后进行一系列分析。肿瘤组织在解剖显微镜下检查并计数,组织病理学由专业病理学家鉴定。
2.5 MTT分析、Transwell测定和集落形成
用BRB花色苷处理HCT116和LoVo细胞48小时,通过溴化3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑鎓(MTT,Sigma-Aldrich,Inc。)测定细胞生长(17),而细胞迁移和侵袭分别通过Transwell和集落形成测定来确定。对于transwell测定,将细胞接种在transwell的顶部室中,而将无血清培养基加入到底部室中。24小时后,用4%PFA固定transwell插入膜,迁移到膜下侧的细胞用DAPI染色(北京鼎国昌盛生物技术有限公司)。对于每个插入物,随机选择10个区域,并使用Image-Pro Plus 6.0对细胞数量进行定量分析。为了计算迁移指数,将每个处理组的细胞数量与对照组的细胞数量标准化处理。对于集落形成测定,将细胞以200个细胞/孔的密度接种在6孔板中并生长2周。出现的细胞集落用磷酸盐缓冲盐水洗涤,用甲醇固定,并用0.5%吉姆萨染色。在倒置显微镜下计数每孔的菌落数。
2.6肠内容物中DNA的提取
每组三只小鼠的肠内容物DNA使用粪便DNA提取试剂盒(北京Tiangen生物技术)进行提取。根据制造商的说明书,然后进行16S rRNA分析。
2.7 T-RFL分析和T-RFs检测
使用6-羧基荧光素(FAM)标记的正向引物(5'-FAM-AGAGTTTGATCCTGGCTCAG-3')和未标记的反向引物1525r(5'-AAGGAGGTGWTCCARCC-3' )分析16S rRNA。通过1%琼脂糖凝胶电泳评估PCR产物的预期大小(1500bp)和纯度。然后使用PCR纯化试剂盒(Sangon Biotech Co.,Ltd.,Shanghai,China)纯化PCR产物。将纯化的PCR产物用限制性酶HhaI,HaeIII和MspI消化,然后送至上海生工生物技术有限公司进行毛细管测序。主要的T-RFs是通过计算机模拟来确定的,这是通过在线系统在http://mica.ibest.uidaho.edu进行的,以分析微生物群落的组成(18)。
2.8 RNA提取和定量RT-PCR
从每只小鼠的肠上皮细胞中提取总RNA(每组3只小鼠)。 通过用分光光度计(Thermo Scientific,NanoDrop-2000c,Gene Co.,Ltd.,Hong Kong,china)测量的UV吸光度测定RNA的质量和数量。如文献(13)所述使用ABI Prism 7500-HT序列检测系统(96孔)进行定量实时PCR分析。如附表1A所示设计和合成以下用于小鼠细胞因子分析的引物。β-actin基因被用作内部对照。还通过qRT-PCR测量了不同处理组小鼠肠道内真杆菌(Eacacterium rectale)(19),柔嫩梭菌群(Faecalibacterium prausnitzii)(19),乳酸杆菌群(Lactobacillus group)(20),脱硫弧菌(Desulfovibriosp)(21)和肠球菌(Enterococcus spp)(22)。如附表1B所示,使用以下引物,并使用16SrRNA作为内部对照。
2.9 DNA提取和甲基化特异性PCR
利用分光光度法测定从三只小鼠以及HCT116和LoVo细胞分离的肠上皮细胞提取的DNA的质量和数量。根据制造商的说明,使用DNA甲基化试剂盒(ComWin Biotech Co.,Ltd.,Beijing,China)对分离的DNA进行亚硫酸氢钠修饰。然后使用甲基化特异性PCR(MSP)分析甲基化。样品含有2×GC缓冲液Ⅱ、1.25mM dNTP混合物、0.5mM每种引物和0.5U LA Taq(Takara bio inc,USA)、50ng亚硫酸氢盐处理的DNA和25μl终体积的水。第一轮循环中PCR反应包括95℃热启动5分钟,然后在95℃45秒,65℃45秒和72℃1分钟的,在接下来的每个循环中退火温度降低1°C。10个循环后,条件变为95℃45秒,55℃45秒,72℃1分钟,再35个循环,然后在72℃延伸10分钟。利用2%琼脂糖凝胶电泳检查PCR产物并在UV-照射下观察。如附表2所示使用以下引物。
2.10免疫印迹分析
如前所述,从肠上皮细胞中提取蛋白质(13)。将总蛋白(40μg)溶解在12%SDS-聚丙烯酰胺凝胶中,然后转移到硝酸纤维素膜上。 将下列一抗用于免疫印迹:抗p-JNK,抗p-STAT3,抗Bcl2,抗Bax,抗CDK4,抗CyclinD1,抗cMyc,抗DNMT1,抗DNMT3A,抗DNMT3B和抗β-actin(Bio Basic Inc.,Canada)。用5%BSA(Sangon Biotech,上海有限公司)在室温封闭2小时后,将膜与适当浓度的一抗在4℃温育过夜。然后将其洗涤并与适当的二抗在室温下孵育2小时,随后使用增强的化学发光技术(Amersham Life Science,USA)进行检测。
2.11统计学分析
利用用SPSS软件(SPSS16; Beijing Stats Data Mining,Beijing,China)对数据进行分析单因素方差分析(ANOVA)及事后检验。
3. 结果
3.1 BRB花色苷降低体内肿瘤多样性和体外增殖、迁移以及集落形成
BRB花色苷的化学预防作用首先通过检测它们对用AOM / DSS处理小鼠的肿瘤发生的抑制作用来证实。所有用不含BRB花色苷的饮物饲喂的AOM / DSS处理小鼠在结肠和直肠中产生高密度肿瘤(>30个肿瘤),而用含有BRB花色苷(LBA或MBA)饮食喂养的AOM / DSS处理小鼠仅平均出现3个或2.25个肿瘤(图1A)。在AOM / DSS处理组中,给予含有BRB花色苷的饮食干预后,肿瘤发生率也轻微降低。没有给予BRB花色苷组肿瘤发生率为100%(所有11只小鼠)、LBA组肿瘤发生率为72.7%(8/11)、MBA组肿瘤发生率为81.8%(9/11),但在这两种情况下减少不显著(图1B)。
为了进一步证实BRB花色苷对肿瘤发生的抑制作用,将HCT116及LoVo细胞与BRB花色苷一起孵育,然后通过MTT测定法测定细胞活性。25或50μg/ ml浓度的BRB花色苷显著降低两种细胞系的活性(图1C),同时我们也注意到花色苷对LoVo细胞活性的降低不是剂量依赖的。在HCT116细胞,我们观察到用25μg/ ml BRB花色苷处理细胞后细胞活性的降低伴随着迁移细胞减少76%、集落形成减少66%,50μg/ ml BRB花色苷处理后在迁移细胞和集落形成分别减少92%和46%(图1D)。用两种不同浓度的BRB花色苷处理LoVo细胞观察到迁移细胞和集落形成出现类似的减少(图1E),并且较低剂量花色苷处理时对集落形成的抑制大于高剂量。结果提示,用BRB花色苷处理这些癌细胞可以抑制它们的侵袭性和集落形成能力。
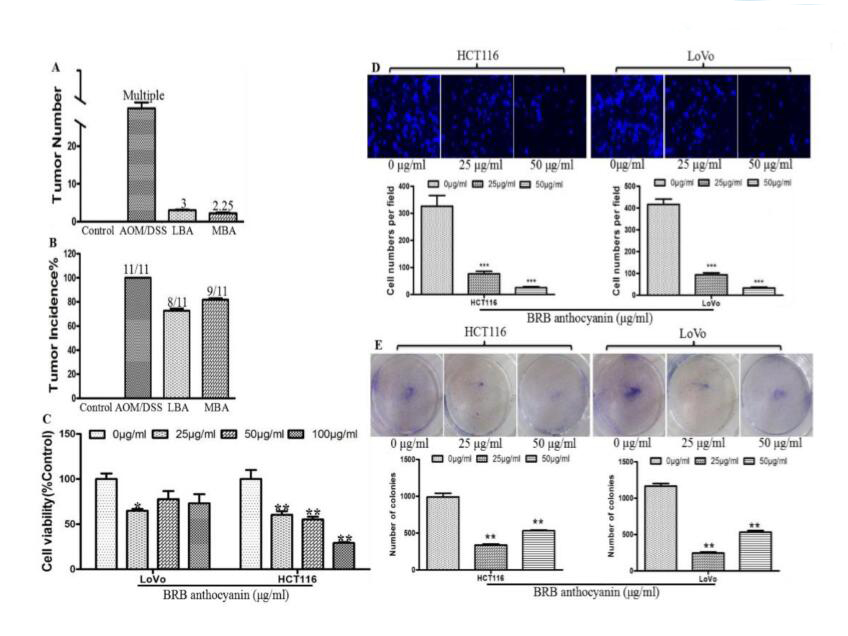
图1 BRB花色苷对体内肿瘤多样性、体外增殖、迁移和集落形成的影响
3.2 T-RFLP分析BRB花色苷对肠道微生物组成的影响
通过T-RFLP评估所有实验组中肠道微生物组成的变化。各组小鼠肠道微生物群落丰富度、多样性和均匀度指数均无显著差异,提示BRB花色苷对肠道菌群的影响可能不大(图2A)。然而,聚类分析表明AOM / DSS处理小鼠给予BRB花色苷和健康小鼠的肠道菌群相似性更高,这两者与未给予BRB花色苷的AOM / DSS处理组显著不同(图2B)。 此外,给予AOM / DSS处理小鼠BRB花色苷,促进了包括丁酸生产菌和奈瑟球菌在内的有益肠道菌群的生长,而肠道致病菌包括弯曲杆菌、幽门螺杆菌、类杆菌和普雷沃氏菌的生长受到抑制(图2C-D)。

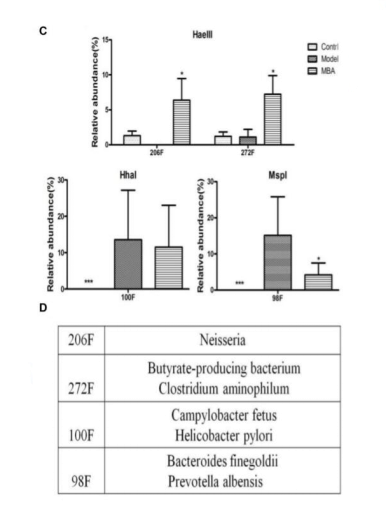
图2 BRB花色苷对肠道菌群组成影响的T-RFLP分析
3.3 BRB花色苷对粪便中不同菌株变化的影响
通过qRT-PCR进一步定量肠内微生物群落的变化,以检查BRB花色苷对个体细菌种群的影响。与健康组小鼠相比,AOM / DSS处理组小鼠肠道细菌包括直肠真杆菌、柔嫩梭菌和乳酸杆菌的数量显著减少,但是,当这些小鼠被给予BRB 花色苷 (MBA组)处理时,这种趋势发生逆转,导致这些细菌数量显著增加(图3)。在AOM / DSS处理组中脱硫弧菌和肠球菌的数量显著增加,而当用MBA干预AOM / DSS处理组小鼠时,脱硫弧菌和肠球菌的数量显著减少。
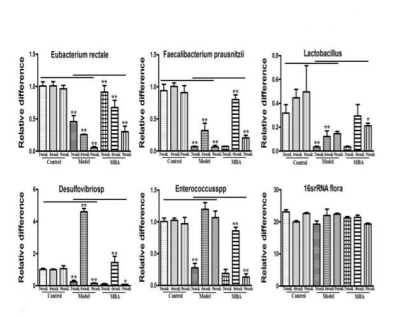
图3 BRB花色苷引起的肠道中不同菌株的变化
3.4 BRB花色苷改善AOM / DSS引起的肠道炎症
为了进一步探讨BRB花色苷对AOM / DSS处理小鼠慢性炎症性引发的肠源性肿瘤发生的影响,采用免疫细胞化学方法检测细胞因子IL-1β、IL-6、IL-10、COX2和TNF-α。与AOM / DSS处理组相比,高剂量BRB花色苷(MBA组)的炎症细胞因子IL-1β、IL-6、COX2和TNF-α水平显著降低(P<0.05)。在较低剂量BRB花色苷(LBA组)饮食AOM / SDD处理小鼠中也观察到IL-1β、IL-6、COX2和TNF-α的表达水平降低,而不是抗炎细胞因子IL-10。提示补充BRB花色苷可显著降低AOM / DSS处理小鼠肠道炎性细胞因子的升高。(图4)
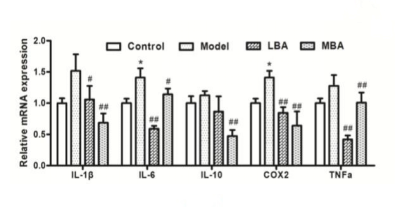
图4 肠内多种细胞因子的mRNA水平变化
3.5 BRB花色苷下调DNA甲基转移酶(DNMTs)的表达并且去甲基化SPFR2基因的低甲基化启动子
无论是在实验室还是临床实践,许多证据都表明肠道菌群与DNA甲基化之间具有密切关系。AOM/DSS诱导结直肠癌形成过程中DNA甲基化改变所显示的与肠道微生物紊乱有关的表观遗传状态的变化也在本研究中进行了研究。哺乳动物细胞中的DNA甲基化受高度相关的DNA甲基转移酶家族(DNMT1,DNMT3A和DNMT3B)的调控。用Western印迹分析DNMTs在肠上皮细胞中的表达。饲喂LBA或MBA的小鼠中DNMT1的水平降低,MBA对DNMT1表达具有显著的影响。Ten-eleven易位(TET)蛋白是介导活性DNA去甲基化的Fe(II)-和2-酮戊二酸(2OG)- 依赖性双加氧酶。为了进一步研究去甲基化过程是被动或主动,测定了BRB花色苷对TET的影响。在给予BRB花色苷小鼠的肠上皮细胞中没有观察到TET1,TET2和TET3 mRNA水平的变化(图5B),表明BRB花色苷调节去甲基化是被动过程。
以往的研究表明Wnt信号传导途径的异常活化可能是由于其上游抑制剂被高甲基化(23-24)。 使用MSP,在HCT116和LoVo细胞体外和体内定量SFRP2和WIF1启动子(Wnt抑制因子1)的甲基化状态,两者都是Wnt信号传导途径的拮抗剂(图5C-5E)。用浓度为25或50μg/ ml的BRB花色苷处理HCT116和LoVo细胞。使用的阳性对照是DNA甲基转移酶抑制剂5-Aza(5-aza-2'-deoxycytidine)。两种细胞系中SFRP2启动子的甲基化水平均降低(图5C-D)。而在相同条件下,BRB花色苷对HCT116和LoVo细胞中WIF1启动子甲基化的影响不明显。
BRB花色苷对SFRP2甲基化的影响还通过检测小鼠分离肠上皮细胞中SPRF2甲基化的程度来评估。AOM/DSS处理小鼠肠上皮细胞中SPFR2的CpG区高甲基化,但当给予BRB花色苷饮食时,这种甲基化减少,与HCT116细胞获得的结果一致(图5E)。此外,BRB花色苷饮食也上调SFRP2蛋白表达(图5F)。数据表明BRB花色苷可能导致SPFR2基因低甲基化启动子的去甲基化,而不是WIF1基因的体内和体外去甲基化。
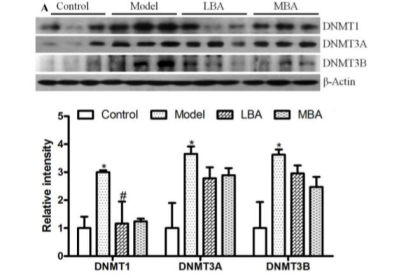
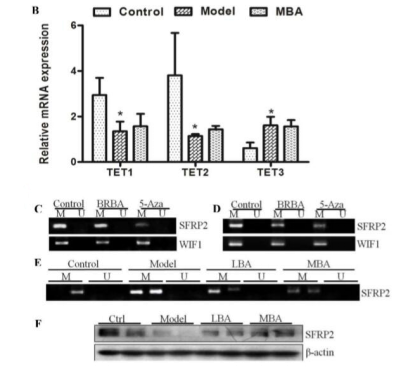
图5 DNA甲基转移酶的变化分析
3.6 BRB花色苷下调肿瘤发生相关基因的表达
AOM / DSS诱导的结直肠癌是一个以多个阶段为特征的过程,涉及多个基因的相互作用。为了进一步了解BRB花色苷对AOM / DSS致癌作用的保护机制,研究了与肿瘤发生有关的各种基因的蛋白水平。BRB花色苷显著抑制了AOM / DSS诱导的肠上皮细胞p-STAT3(信号转导体和转录激活因子3,STAT3)水平的降低。此外,参与β-catenin信号通路的基因(包括p-JNK,Bcl2,CDK4,CyclinD1和c-Myc)可促进肿瘤增殖并抑制细胞凋亡,这些基因的表达被BRB花色苷显著下调。BRB花色苷上调凋亡诱导因子Bax的表达。(图6)
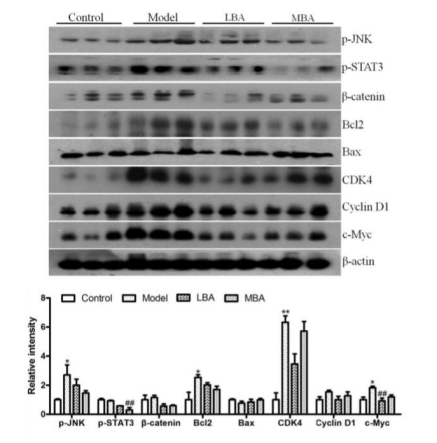
图6 BRB花色苷对肠上皮细胞的多种基因及结直肠癌发生发展的影响
4. 讨论
越来越多的证据表明,肠道微生物的复杂群体与结直肠癌的发展密切相关[25]。益生菌在降低胃肠道炎症和预防结直肠癌方面的作用已经被证实,但是它们在抑制肿瘤生长中的免疫调节作用和机制尚不清楚。目前的研究旨在探讨肠道微生物在BRB花色苷对结直肠癌化学预防过程中的作用。我们的研究结果清楚地表明,给予AOM / DSS处理小鼠BRB花色苷饮食可以显著改善破坏的肠道微生物群,而肠道菌群破坏会诱导肿瘤表观遗传、遗传改变或使炎症加剧。通过增加益生菌和减少病原菌数量来调节肠道微生物群的组成,BRB花色苷在AOM / DSS诱导的大肠癌小鼠模型中发挥了化学预防作用。
肠道微生物失衡表现为微生物多样性整体下降,包括柔嫩梭菌在内的厚壁菌门丰度下降同时伴随拟杆菌增加,将导致慢性肠道炎症并增加结直肠癌的易感性[26]。在这项研究中,我们发现在AOM / DSS处理小鼠中观察到100%的肿瘤发生率,并且BRB可以显著减小肿瘤(图1A),与先前报道的APC1638和MUC2小鼠一致(13),在减少肿瘤发病率方面,BRB花色苷所起的作用没有以前的研究那么显著。与以前的研究相比,动物被给予BRB花色苷的持续时间、遗传背景、饮食组成和动物饲养的物理环境的差异可能导致了当前研究中获得的不同结果。目前令人印象深刻的发现是在为期12周的实验中除了在AOM / DSS诱导的动物中检测到100%肿瘤发生率之外,致病性细菌(包括嗜血杆菌,大肠杆菌和肠道沙门氏菌)的数量也显著增加。真细菌(Eubacteria)与CRC有显著的相关性。真细菌是一种能够从宿主的饮食中获取能量的细菌(27),在结直肠癌患者的肠道菌群中发现了它们的减少(28)。相反,在CRC患者中通常富含的变形菌通常被认为是具有潜在致病特征的肠道共生菌(29)。厚壁菌门是没有给予BRB花色苷的AOM/DSS处理小鼠肠道内容物中最主要的细菌,其中致病性亚群,嗜血杆菌、大肠杆菌和肠道沙门氏菌更为丰富。与没有给予BRB花色苷的AOM/DSS处理小鼠相比,在给予BRB花色苷的AOM / DSS处理小鼠的肠内容物中这些细菌数目的显著减少(图3)。丁酸盐是一种由未被吸收的碳水化合物细菌发酵产生的短链脂肪酸,为结肠上皮细胞提供能量、促进上皮细胞分化、改善炎症、加速结肠组织的修复(30)。肠道真细菌和柔嫩梭菌是肠道中两种主要的产丁酸菌(31)。乳酸杆菌可以改善结肠癌的发生,抑制癌前病变,减少肿瘤负荷和大小(32)。目前的研究结果表明,膳食BRB花色苷补充剂的有益调节作用确实可以增强真细菌、柔嫩梭菌和乳酸杆菌的生长,并抑制脱硫弧菌和肠球菌的生长。
此外,AOM/DSS处理小鼠的炎性细胞因子,包括IL-1β、IL-6、COX2和TNF-α的释放显著增加。BRB花色苷可降低上述四种炎性细胞因子的水平。最显著的是TNF-α和IL-6,这与我们先前在Muc2-/-小鼠中的发现一致(13)。在炎症期间,结肠上皮细胞和免疫细胞中的NF-κB水平增加,导致促炎细胞因子增加(33)。然后这些事件会增加DNA甲基转移酶(DNMTs)的活性,DNMTs可以通过启动子的甲基化沉默一部分肿瘤抑制基因。DNMT1在哺乳动物细胞中维持DNA甲基转移酶是必不可少的,并且负责在细胞周期的S期期间精确复制基因组DNA甲基化模式(34)。相反,DNA的从头甲基化是由DNMT3A和DNMT3B介导的,它们既具有维持性又具有从头DNA甲基化活性(35)。所有三种DNMT都在肿瘤组织中过度表达(36)。Wang等人早期(37-40)研究了BRB花色苷对人结肠癌细胞、溃疡性结肠炎(UC)相关小鼠模型和人结直肠癌患者DNA甲基化的影响,结果表明BRB花色苷不仅能够抑制DNMT3B和DNMT1在人结直肠癌细胞以及溃疡性结肠炎相关的小鼠中的表达,而且当人类大肠癌患者平均服用BRBs 4周后,能够降低活检标本中DNMT1蛋白水平。同时,他们的数据也表明,BRB花色苷可以与DNMT3B和DNMT1在HCT116细胞中共定位。我们的数据提供了在这种AOM / DSS诱导的溃疡性结肠炎相关的结直肠癌小鼠模型中BRB花色苷对DNMT1抑制作用的额外证据。DNMT3B也表现出减少的模式,但是,这并不显著。
已知所有三个TET基因都发生了突变,并且在mRNA水平上表现出表达降低,并且相应的蛋白质在不同癌症类型(包括结直肠癌)中活性受损(41)。我们的数据表明,BRB花色苷通过被动模式调节小鼠结直肠癌细胞中SFRP2的甲基化,因为在BRB花青素干预的AOM / DSS处理小鼠肠上皮细胞观察到的TET1,TET2和TET3的mRNA水平实际上没有变化(图5B)。Wnt通路的负调控因子在溃疡性结肠炎中经常被甲基化,导致通路失调,并可能导致结直肠癌[23]。由于假定WNT活性的抑制,因此推测SFRP起到肿瘤抑制剂的作用。已经表明SFRP的表达受表观遗传沉默的影响(24,42)。BRBs花色苷介导的SFRP2基因去甲基化导致其在HCT116和LoVo人结直肠癌细胞和小鼠肠上皮细胞中表达水平增加(图5F),这与Wang等人以往报道的人结肠癌样本中的发现一致(39)。同时这也导致下游因子β-catenin、CDK4、CyclinD1、c-Myc水平下降。这些结果表明BRB花色苷可以通过调节参与抑制结肠炎症的基因启动子的高甲基化来抑制结肠溃疡。
已经报道了由STAT3活性介导的白细胞介素6或组成性活化的STAT3突变体触发的CRC细胞增殖(43)。在CRC患者中,与健康个体相比,STAT3和p-STAT3的水平显著升高,可能伴随着Bcl-xl水平的增加,也可以促进肿瘤增殖[44]。STAT3可导致恶性肿瘤,下调其表达被认为是治疗CRC的潜在方法。抑制小鼠IL-6/STAT3反式信号传导可以有效地抑制结肠癌的生长(29)。在BRB花色苷干预小鼠中观察到p-STAT3、p-JNK1和Bcl2的表达显著降低,Bax表达增加(图6)表明抑制STAT3-信号传导可能确实是BRB花色苷调节肠道微生物群、调节炎症和最终预防CRC形成和发展的关键因素。这一发现与之前的报道是一致的,肠道菌群可促进STAT3信号传导途径,加速CRC小鼠肿瘤生长 [45]。
综上所述,在AOM/DSS诱导的溃疡性结肠炎相关性大肠癌的发病过程中,肠道微生物的破坏可能是一个早期事件,可能引发肠上皮细胞发生炎症和表观遗传学改变,最终导致肿瘤的形成和发展。BRB花色苷可能通过维持保护性细菌的生长而不是致病细菌的生长以及通过调节肠道微生物群的组成和共生体来充当有效的益生元。因此,BRB花色苷的化学预防效果可以集中于由肠道菌群稳态失衡引起的SFRP2炎症和异常表观遗传状态的调节。
(声明:本文由福山驱动翻译,转载请注明来源)
原文标题:
Chen L, Jiang B, Zhong C, et al. Chemoprevention of colorectal cancer by black raspberry anthocyanins involved the modulation of gut microbiota and SFRP2 demethylation[J]. Carcinogenesis, 2018.
本文来源于Carcinogenesis杂志,如有侵权,请联系我们删除。