






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
萝卜硫素与吡哆胺的补充使2型糖尿病中的内皮功能紊乱恢复正常
发表于:2019-05-13 作者:admin 来源:本站 点击量:14570
摘要
在本研究中,我们把吡哆胺(PM)和/或萝卜硫素(SFN)作为治疗干预措施,用于确定NFE2相关因子2(Nrf2)的激动剂是否可以同晚期糖基化终产物(AGE)形成抑制剂一起使用,达到减轻2型糖尿病患者的氧化应激和改善其内皮功能障碍。Goto Kakizaki(GK)大鼠,一种非肥胖型2型糖尿病动物模型,在8周内接受或不接受PM和/或SFN治疗,并与年龄匹配的Wistar大鼠进行比较。在治疗结束时,评估孤立主动脉和肠系膜动脉中一氧化氮(NO)依赖性和非依赖性的血管舒张。还评估了代谢概况、NO生物利用度和血管氧化应激、AGE和Nrf2水平。糖尿病GK大鼠的Nrf2水平显著降低,同时氧化应激和内皮功能障碍水平也较高。PM和SFN作为单一疗法能够显著改善主动脉和肠系膜动脉的内皮功能障碍,降低血管氧化损伤、AGE和HbA1c水平。此外,在GK大鼠中已证明,SFN+PM能更有效地降低全身游离脂肪酸水平,使内皮功能、NO生物利用度和糖化作用正常化。Nrf2激动剂可与AGE抑制剂和交联形成抑制剂联用来治疗2型糖尿病内皮功能障碍。
2型糖尿病与氧化应激水平升高和糖基化改变血管功能有关,因此被认为是心血管疾病发展的主要危险因素。许多的糖尿病并发症都与高血糖和活性氧物种(ROS)生成增加有关,其导致了内皮功能紊乱。对血管舒张剂(如乙酰胆碱)的内皮依赖性舒张功能受损是1型和2型糖尿病实验模型中导管和阻力动脉的共同特征。我们先前发现糖尿病GK大鼠的氧化应激和糖基化增加,而这种变化导向内皮功能障碍。
糖尿病的治疗进展并不能有效地减少糖尿病相关的血管并发症。考虑到ROS在内皮功能中的关键作用,人们已经作出了相当大的努力来探索降低血管系统中ROS的治疗方法。核因子E2(红系衍生2)相关因子-2(Nrf2)是一种转录因子,在细胞对氧化应激的保护中起着关键作用。Nrf2被称为抗氧化剂反应的“主调节器”。通过Nrf2/抗氧化反应元件途径激动剂诱导内源性抗氧化酶可能是获得足够水平的抗氧化剂和减少氧化应激的一种有趣方法。膳食中的NRF2激动剂,如十字花科蔬菜中发现的异硫氰酸盐萝卜硫素(SFN),可以增加抗氧化防御,降低血压,抑制肾脏内促炎信号通路,并预防高血糖引起的内皮细胞代谢紊乱。在糖尿病模型中,Nrf2的激活降低了野生型小鼠的血糖水平,减少了糖尿病相关肾病,但在Nrf2缺乏的小鼠中则没有。因此,Nrf2诱导剂在血管疾病中的治疗潜力巨大,特别是与长期2型糖尿病有关的血管疾病。
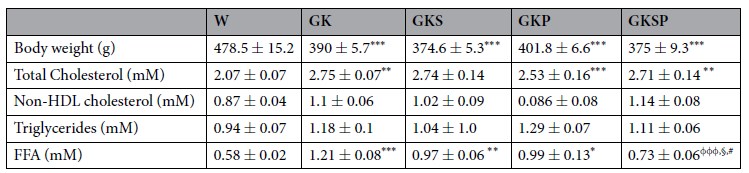
表1 8个月大的非糖尿病Wistar大鼠(W)、糖尿病Goto-Kakizaki大鼠(GK)、接受萝卜硫素处理的GK大鼠(GKS)、接受吡哆胺处理的GK大鼠(GKP)和接受萝卜硫素与吡哆胺处理的GK大鼠(GKSP)的体重和血脂水平。数据表示为平均值±SE(n=12每组动物)。*P<0.05,**P<0.01,***P<0.001 vs W大鼠。与GK大鼠相比,φφφP<0.001。§P<0.05 vs GKS大鼠。#P<0.05 vs GKP大鼠。
此外,在糖尿病情况下的晚期糖基化加速导致血管并发症。几位作者已经证明,抑制晚期糖基化终产物(AGE)形成可能是改善糖尿病血管并发症的一种治疗策略。吡哆胺(PM)是维生素B6的一种天然衍生物,已被证明是一种有效的蛋白质糖化和脂质氧化抑制剂。在体外和体内,PM均能抑制AGE的形成,对糖尿病肾病和视网膜病变的治疗具有潜在的益处。
萝卜硫素和吡哆胺有不同的作用机制。因此,我们假设单独或联合使用SFN、PM对2型糖尿病模型Goto-Kakizaki(GK)大鼠的内皮依赖性血管反应性、氧化应激和代谢参数具有有益影响。在糖尿病进展的同时,对脂质分布、氧化应激、糖基化和一氧化氮(NO)生物利用度进行了评价。此外,还评估了主动脉和肠系膜动脉对乙酰胆碱(ACH)和硝普钠(SNP)的内皮依赖性和非依赖性血管敏感性。与Wistar大鼠相比,8个月大的GK大鼠表现出长期糖尿病表型,导管和阻力动脉内皮功能障碍,以及氧化应激增加。单用SFN和PM可以改善孤立动脉中的NO依赖性血管舒张。顺带一提,SFN和PM通过一种机制使主动脉和肠系膜动脉的内皮功能正常化,该机制包括NO生物利用度的增加,以及氧化应激和AGE水平的降低。总之,这些研究表明,针对引起内皮功能障碍的不同机制,抑制AGE形成(PM)和促进抗氧化防御系统(SFN)增强应是对抗糖尿病大血管并发症的有用工具,因其同时作用于最终导致血管损伤的不同机制。
结论
动物特征。我们实验中使用的糖尿病大鼠在研究开始时表现出类似的空腹血糖水平。治疗前的GK大鼠血糖(GK6M)明显高于年龄匹配的非糖尿病大鼠(图1补充)。GK大鼠的体重明显低于年龄匹配的Wistar大鼠,不同的治疗方法并没有显著改变这个参数(表1)。在腹膜内葡萄糖耐量试验(IPGTT)中,GK大鼠与Wistar大鼠相比表现出明显的葡萄糖不耐(图1A)。与Wistar大鼠相比,GK大鼠的空腹血糖、葡萄糖曲线下面积(AUC)、HbA1c、总胆固醇和游离脂肪酸(FFA)升高(图1B、C、D、表1)。如前所述,GK大鼠表现出正常的非HDL胆固醇和甘油三酯水平(表1)。单用SFN或PM治疗并没有显著改变糖耐量和脂质分布评估(图1A、C,表1)。同时,PM显著降低了空腹血糖和HbA1c水平(图1B、D)。事实上,所有的治疗方法都能有效降低GK大鼠的HbA1c水平(图1D)。此外,使用SFN和PM治疗8周可有效降低糖尿病大鼠的血液中FFA浓度(表1),并显著改善葡萄糖耐受性(图1A、C)。
NO依赖性血管舒张。8个月大的GK大鼠与年龄匹配的Wistar大鼠相比,苯肾上腺素预收缩后的主动脉环响应乙酰胆碱(ACh)引起的内皮介导血管舒张能力受损(图2A),但对硝普钠(SNP)引起的非内皮依赖性舒张在这两个物种中相似(图3A)。事实上,在糖尿病大鼠中,苯肾上腺素预收缩主动脉环对乙酰胆碱引起的内皮介导血管最大舒张值降低了49%(图2A)。在GK大鼠和Wistar大鼠中,用NOS抑制剂N-硝基-L-精氨酸甲酯(L-NAME)和环氧合酶抑制剂吲哚美辛预孵化后的动脉环几乎完全消除了Ach的松弛作用(图2补充)。在我们的实验条件下,其他血管扩张剂的残余成分约为15%。在主动脉中,没有观察到不同组间SNP量效曲线在最大舒张值上的差异(图3A)。所有GK治疗组的最大舒张值显著改善,对Ach的血管反应性有所改善(图2A,表2)。用SFN、PM或SFN+PM治疗可显著改善糖尿病大鼠主动脉内皮依赖性血管舒张(图2A,表2)。这种血管舒张对吲哚美辛不敏感,并被L-NAME阻断,表明其机制依赖于内源性NO释放(图2补充)。此外,在所有GK治疗组中观察到SNP敏感性增加(图3A,表2)。在肠系膜动脉中,糖尿病大鼠的苯肾上腺素预收缩后动脉环响应Ach引起的内皮介导的最大舒张值降低了36%(图2B)。此外,糖尿病大鼠响应SNP引起的最大舒张值下降了39%(图3B)。用SFN、PM或SFN+PM治疗能够改善内皮依赖性血管扩张(图2B,表3)。类似地,这种作用对吲哚美辛不敏感,并被L-NAME抑制(图2补充)。表2和表3总结了最大舒张值和EC50值的详细数据。这些结果表明,单用SFN或PM治疗可改善糖尿病GK大鼠主动脉和肠系膜动脉的内皮依赖性血管舒张。与此相关,这些疗法能够使内皮介导的舒张正常化,并提高主动脉和肠系膜动脉的SNP敏感性(图2、3)。
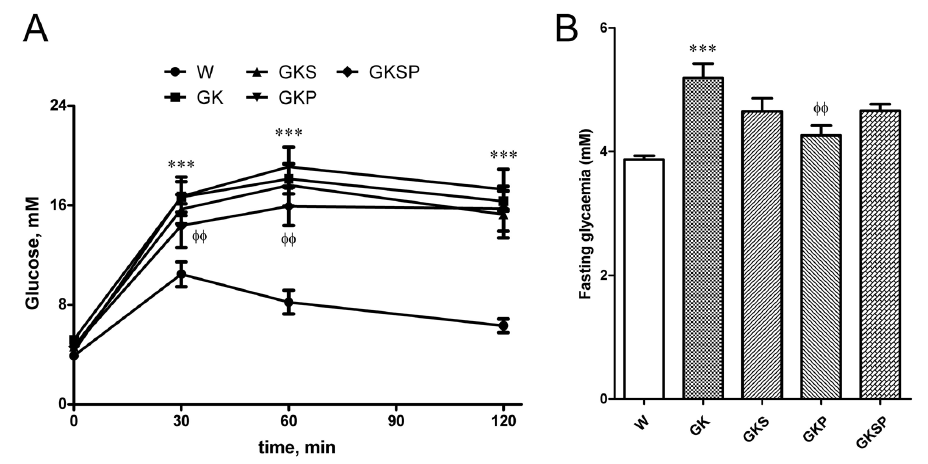

图1 与非糖尿病Wistar(W)大鼠相比,8个月大的糖尿病Goto-Kakizaki(GK)大鼠萝卜硫素和吡哆胺处理的效果,腹腔葡萄糖耐量试验(IPGTT;A)、空腹血糖(B)、葡萄糖曲线下面积(AUC;C)和糖化血红蛋白(HbA1c)水平(D)。C)计算IPGTT曲线的AUC,以测量葡萄糖耐量受损的程度。数据表示为平均值±SE(n=12)。***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组。
血管壁的氧化应激。我们确定了SFN、PM或两者对糖尿病血管系统氧化应激的潜在影响。有趣的是,糖尿病导致糖尿病大动脉中的超氧化物生成量增加了3倍(p<0.001;图4B,K)。与GK大鼠相比,用SFN、PM和SFN+PM治疗的糖尿病大鼠主动脉中二氢乙啶(DHE,ROS荧光探针)的密度显著降低(图4C、D、E、K)。此外,糖尿病GK大鼠肠系膜动脉中的免疫反应性硝基酪氨酸(蛋白质氧化生物标志物)水平也升高(p<0.001;图4G,L),并且用SFN、PM或两者治疗后,这些水平显著降低(图4H,I,J,L)。因此,所有疗法都能显著降低糖尿病GK大鼠主动脉和肠系膜动脉的血管氧化损伤。相应地,GK大鼠尿中8-羟基-2’-脱氧鸟苷(8-OHdG,DNA氧化损伤生物标志物)的水平明显高于年龄匹配的Wistar大鼠(图4M)。对GK大鼠使用SFN、PM或两者治疗8周后,8-OHdG水平显著降低(图4M)。
主动脉中的肺血管紧张素和总血管紧张素。血管舒张剂激活磷蛋白(VASP)在Ser239的磷酸化被证明是生物活性NO和NO-cGMP蛋白激酶G信号通路活性的指标。为了确定NO生物利用度,我们测量了主动脉中磷酸化血管舒张剂激活磷蛋白(pVASP)和总VASP。GK大鼠的主动脉pVASP/tVASP比率明显低于Wistar大鼠(图5A,B)。在糖尿病GK大鼠(GKS,GKP;GKSP;图5A,B)中,用SFN、PM或两者治疗可显著提高pVASP/TVSP比率。在肠系膜动脉中观察到类似的情况(数据未列出)。
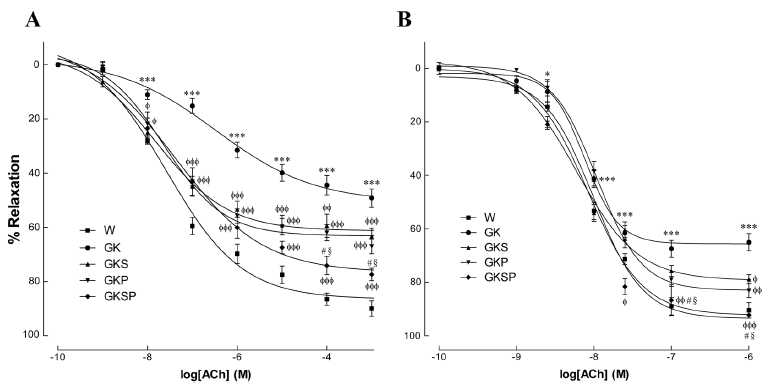 图2 与非糖尿病Wistar大鼠(W)相比,萝卜硫素和吡哆胺处理对GK大鼠主动脉(A)和肠系膜动脉(B)乙酰胆碱舒张反应的影响。数据表示为平均值±SE(n=12)。*P<0.05,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组;§P<0.05 vs GKS组;#P<0.05 vs GKP组。
图2 与非糖尿病Wistar大鼠(W)相比,萝卜硫素和吡哆胺处理对GK大鼠主动脉(A)和肠系膜动脉(B)乙酰胆碱舒张反应的影响。数据表示为平均值±SE(n=12)。*P<0.05,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组;§P<0.05 vs GKS组;#P<0.05 vs GKP组。
 图3 与非糖尿病Wistar大鼠(W)相比,萝卜硫素和吡哆胺处理对GK大鼠主动脉(A)和肠系膜动脉(B)中硝普钠舒张反应的影响。数据表示为平均值±SE(n=12)。*P<0.05,****P<0.001 vs Wistar组;φφP<0.01,φφφP<0.001 vs GK组。
图3 与非糖尿病Wistar大鼠(W)相比,萝卜硫素和吡哆胺处理对GK大鼠主动脉(A)和肠系膜动脉(B)中硝普钠舒张反应的影响。数据表示为平均值±SE(n=12)。*P<0.05,****P<0.001 vs Wistar组;φφP<0.01,φφφP<0.001 vs GK组。
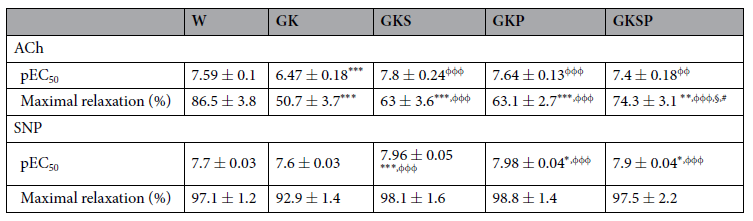 表2 8个月大的不同处理的自发性糖尿病Goto-Kakizaki(GK)大鼠和年龄匹配的非糖尿病Wistar大鼠离体主动脉的最大舒张反应(%)和−logEC50。数据表示为平均值±SE(n=12每组动物)。 pEC50值以激动剂浓度的负对数(−logEC50)表示。*P<0.05,**P<0.01,***P<0.001 vs W大鼠。与GK大鼠相比,φφP<0.01,φφφP<0.001。§P<0.05 vs GKS。 #P<0.05 vs GKP。
表2 8个月大的不同处理的自发性糖尿病Goto-Kakizaki(GK)大鼠和年龄匹配的非糖尿病Wistar大鼠离体主动脉的最大舒张反应(%)和−logEC50。数据表示为平均值±SE(n=12每组动物)。 pEC50值以激动剂浓度的负对数(−logEC50)表示。*P<0.05,**P<0.01,***P<0.001 vs W大鼠。与GK大鼠相比,φφP<0.01,φφφP<0.001。§P<0.05 vs GKS。 #P<0.05 vs GKP。
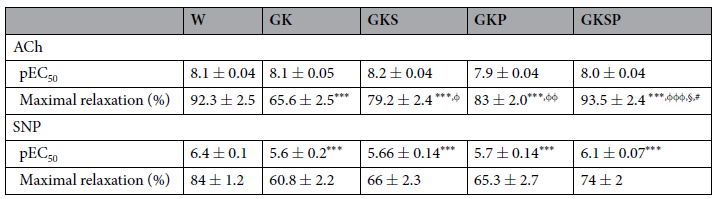 表3 8个月大的自发性糖尿病Goto-Kakizaki(GK)大鼠和匹配的非糖尿病Wistar大鼠肠系膜动脉的最大松弛反应(%)和−logEC50。数据表示为平均值±SE(n=12每组动物)。pEC50值以激动剂浓度的负对数(−logEC50)表示。***P<0.001 vs W大鼠。与GK大鼠相比,φP<0.05,φφP<0.01,φφφP<0.01。§P<0.05 vs GKS。#P<0.05 vs GKP。
表3 8个月大的自发性糖尿病Goto-Kakizaki(GK)大鼠和匹配的非糖尿病Wistar大鼠肠系膜动脉的最大松弛反应(%)和−logEC50。数据表示为平均值±SE(n=12每组动物)。pEC50值以激动剂浓度的负对数(−logEC50)表示。***P<0.001 vs W大鼠。与GK大鼠相比,φP<0.05,φφP<0.01,φφφP<0.01。§P<0.05 vs GKS。#P<0.05 vs GKP。
主动脉和肠系膜动脉中的Nrf2水平。我们随后量化了不同组大鼠动脉中的Nrf2水平。用Western blot检测主动脉中的Nrf2水平,以共聚焦显微镜观察肠系膜动脉中的免疫荧光检来测Nrf2水平。糖尿病大鼠血管中的Nrf2水平明显低于主动脉和肠系膜动脉中年龄匹配的Wistar大鼠(图6I,F)。萝卜硫素治疗组(GKS和GKSP)显著增加了主动脉和肠系膜动脉中的细胞核Nrf2水平(图6J,G)。Nrf2激动剂SFN增加糖尿病GK大鼠Nrf2的核定位,这可能导致抗氧化防御增加,促进血管氧化应激的减少。在糖尿病GK大鼠的主动脉和肠系膜动脉中,SFN显著增加了Nrf2的表达(图6),这表明它同时促进了表达增加和核易位,与其在其他组织中的作用相一致。
主动脉和肠系膜动脉的AGE水平。接下来,我们量化了不同组大鼠动脉中的AGE水平。主动脉和肠系膜动脉的AGE水平用共聚焦显微镜测量。糖尿病大鼠主动脉和肠系膜动脉中的AGE水平明显高于年龄匹配的Wistar大鼠(分别为图7B、K、G、L)。GK治疗组(GKS、GKP和GKSP)主动脉和肠系膜动脉的血管AGE水平显著降低(图7K,L)。值得注意的是,PM治疗组(GKP,GKSP)血管壁具有正常的AGE水平。
讨论
在目前的研究中,萝卜硫素、吡哆胺及其联用对GK大鼠的糖尿病内皮功能障碍表现出明显的治疗潜力。用SFN、PM或两者联合治疗GK大鼠可显著降低主动脉和肠系膜动脉的内皮功能障碍。在这些血管中,SFN、PM或SFN+PM治疗可延缓包括氧化损伤在内的糖尿病内皮功能障碍病理特征的出现。此外,这两种相关疗法都能降低FFA水平,也增加了NO生物利用度,这可以部分解释2型糖尿病长期模型中内皮功能的正常化。
GK大鼠为非肥胖2型糖尿病动物模型,具有轻度高血糖、高胰岛素血症和胰岛素抵抗。长期糖尿病通常与这些动物的胰腺功能丧失有关,其特征也类似于人类疾病进展中的相关特征。该模型与人类2型糖尿病共有包括异常血管反应性在内的多种心血管表型,证明其作为研究糖尿病并发症的模型是合适的。
在此,我们发现8个月大的GK大鼠具有高血糖,显著增加的总胆固醇和循环FFA水平的特征。内皮功能障碍在肠系膜动脉和主动脉动脉中都很明显,其伴随着氧化应激和糖基化作用升高、NO生物利用度下降和Nrf2水平降低。
Nrf2抗氧化功能在血管疾病中可能很重要。SFN是一种在西兰花中发现的Nrf2激动剂,增加抗氧化反应元件相关转录控制下的保护酶的表达,并防止由高血糖引起的内皮细胞代谢功能障碍。在其它研究的基础上,结合西兰花芽苗中SFN的生物利用度,测定了该化合物的浓度。我们估计,在人体饮食可行的生理剂量下,SFN对治疗内皮功能障碍具有潜在的治疗益处。另一方面,我们最近检查了2型糖尿病动物模型的AGE水平,发现血浆和血管壁的AGE水平都在增加。如PM的化合物可以降低高水平的AGE,和这种方法对内皮功能有潜在的有益作用。这两种疗法对大血管疾病的治疗效果尚未被研究过。在本文中,我们研究了在血管水平上,有或没有与PM共同抑制年龄形成的SFN的作用。
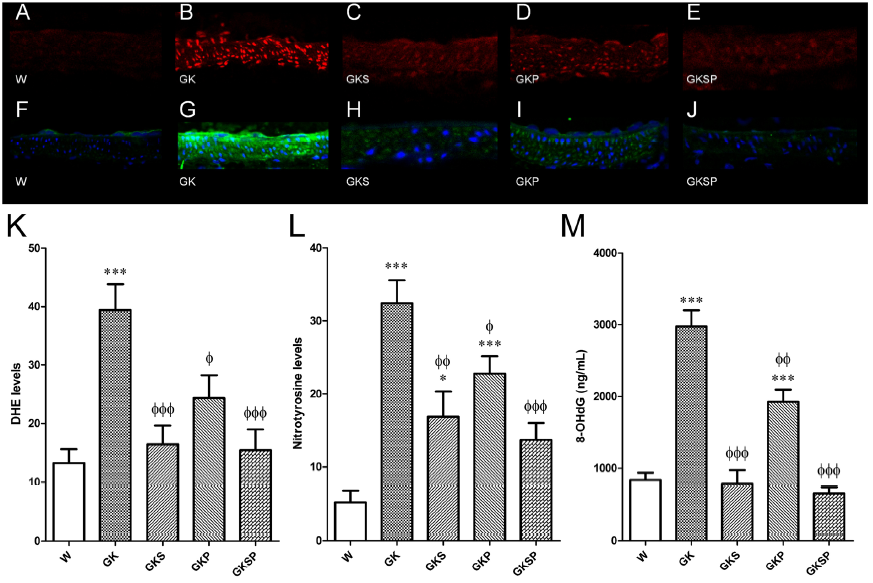
图4 与非糖尿病Wistar(W)大鼠相比,萝卜硫素和吡哆胺处理对GK大鼠全身和血管氧化应激的影响。代表性的DHE染色的主动脉段反映了不同处理(A-E)下的氧气生成。所有的内皮都朝上。在相同的条件下,糖尿病患者GK(B)的荧光明显高于正常血管(Wistar,A)。注意到GK主动脉内皮、内膜和介质中反射O2·−水平的荧光增强。糖尿病GK大鼠服用SFN(GKS,C)、PM(GKP,D)和SFN+PM(GKSP,E)后,DHE荧光降低。面板(K)包含不同动脉组中荧光溴化乙锭信号的量化。具有代表性的肠系膜切片用硝基酪氨酸染色显示非糖尿病Wistar大鼠(F),糖尿病GK大鼠(G),糖尿病GK大鼠SFN(GKS,H)、PM(GKP,I)和SFN+PM(GKSP,J)处理组。面板(L)包含不同动脉组中绿色荧光的量化。(M)不同组大鼠尿8-羟基脱氧鸟苷(8-OHDG)水平。数据表示为平均值±SE(n=12)。*P<0.05,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组。
单用SFN或PM治疗8周并没有显著改善甘油三酯、胆固醇和FFA水平。已经证明,PM可以纠正糖尿病和肥胖大鼠的高脂血症和肾病,可能是通过干扰各种AGE/晚期脂质氧化最终产物(ALE,由脂质氧化形成)的活性羰基中间产物。在不同的动物模型中,更高浓度的PM(400 mg/kg/天)具有这种降脂作用。此外,Nrf2通路具有调节肝脏脂质代谢的功能。Nrf2负性调节许多编码参与脂质生物合成、脂肪酸去饱和与脂肪酸转运的酶的基因。相反,0.5 mg/kg的SFN会损害链脲佐菌素诱导的糖尿病大鼠的肝功能并加重的胆固醇水平。此外,在我们的实验条件下,单用SFN治疗不会改变脂质分布。
SFN+PM治疗对总胆固醇和甘油三酯水平没有影响,但却显著降低了FFA和HbA1c水平。两种联合治疗均能改善葡萄糖耐受性,显著减少葡萄糖曲线下面积。靶向糖基化和抗氧化防御系统的PM和SFN在降低FFA和HbA1c水平方面非常有效。重要的是,SFN和PM治疗对胰岛素水平没有影响,这解释了在治疗糖尿病大鼠中观察到的高血糖。事实上,在所有GK治疗组中,血糖水平明显高于Wistar大鼠。然而,在这些条件下,sfn+pm的血管保护作用是明显的,表明正常血糖的完全恢复不是血管保护的先决条件。
其他研究者先前报道,在非缺陷Nrf2的野生型小鼠中,Nrf2的激活降低了血糖水平,减少了糖尿病相关肾病。而这项研究中,SFN治疗组(GKS)没有促进血糖水平下降。不同的动物模型和实验条件可以解释这些差异。
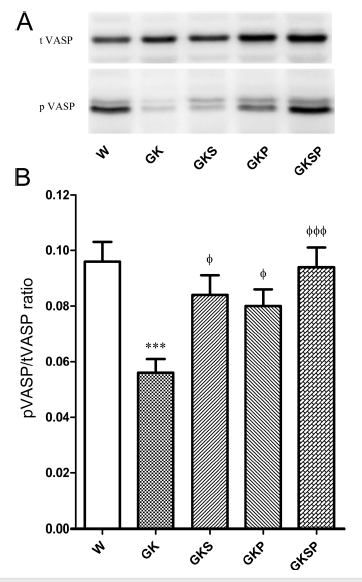 图5 萝卜硫素和吡哆胺处理对pVASP和tVASP表达水平的影响。为了检测NO-cGMP信号激活,评估主动脉中总血管舒张剂刺激的磷酸蛋白(tVASP)和磷酸化(ser239)VASP(pVASP)的表达。用SDS-PAGE法测定主动脉裂解物。(A)不同动脉组的主动脉中总血管紧张素(VASP)和pVASP蛋白水平的典型Western blot条带。(B)pVASP/总VASP比的平均密度测定数据。数据表示为平均值±SE(n=12)。***P<0.001 vs Wistar组;φP<0.05,φφφP<0.001 vs GK组。
图5 萝卜硫素和吡哆胺处理对pVASP和tVASP表达水平的影响。为了检测NO-cGMP信号激活,评估主动脉中总血管舒张剂刺激的磷酸蛋白(tVASP)和磷酸化(ser239)VASP(pVASP)的表达。用SDS-PAGE法测定主动脉裂解物。(A)不同动脉组的主动脉中总血管紧张素(VASP)和pVASP蛋白水平的典型Western blot条带。(B)pVASP/总VASP比的平均密度测定数据。数据表示为平均值±SE(n=12)。***P<0.001 vs Wistar组;φP<0.05,φφφP<0.001 vs GK组。
氧化应激在内皮功能障碍发病机制中的作用已被充分确证。糖尿病增加血管系统中的活性氧,损害抗氧化防御酶,并降低细胞内抗氧化物的水平,创造一个氧化应激增加的环境。在血管模型中,Nrf2通路激动剂通过增加抗氧化/亲电反应元件介导的II相酶和抗氧化酶表达(包括NAD(P)H:醌氧化还原酶-1、血红素氧合酶-1和γ-谷氨酸-半胱氨酸连接酶催化亚单位)来恢复氧化还原平衡。在这项研究中,我们提出的证据表明,SFN和PM治疗在体内具有抗氧化作用:在糖尿病GK大鼠中,SFN、PM或SFN+PM降低了尿8-OHdG和组织中O2•−阴离子以及硝基酪氨酸的积累量。这种抗氧化作用是已观察到的内皮功能障碍正常化机制的重要因素。因此,其他研究也报道了不同条件下SFN对氧化应激的抑制作用。在糖尿病db/db微血管系统中,用SFN靶向Nrf2通路能有效恢复正常的氧化还原平衡和对抵抗小动脉的肌源性反应。此外,通过介导上调Nrf2活性,SFN下调高脂饮食喂养的小鼠主动脉中血管细胞粘附分子-1的表达。我们的数据支持一个模型,在该模型中,通过一种涉及Nrf2依赖性的抗氧化能力下降的机制,主动脉和肠系膜动脉中的氧化应激增加。我们在活体内证明,用SFN靶向Nrf2通路可有效恢复正常的氧化还原平衡,改善糖尿病血管系统中主动脉和肠系膜动脉对乙酰胆碱的内皮反应。这些有益的影响被PM增强,而PM是一种AGE/ALE和其交联形成的抑制剂。
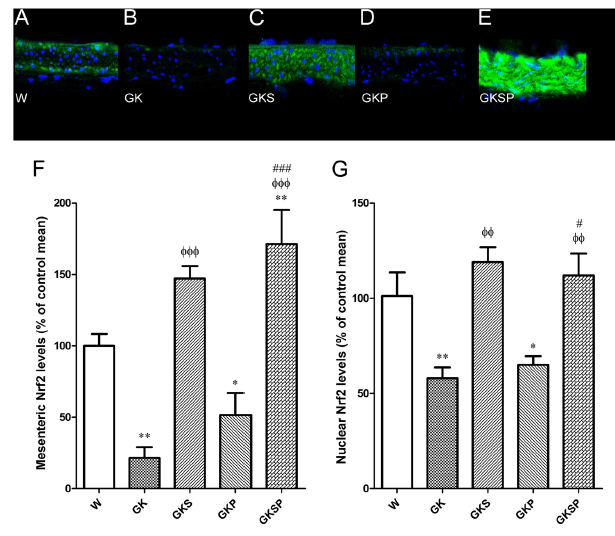

图6 与非糖尿病Wistar大鼠相比,萝卜硫素和吡哆胺对GK大鼠主动脉和肠系膜动脉中Nrf2水平的影响。代表性肠系膜切片显示非糖尿病性Wistar大鼠(A)、糖尿病性GK大鼠(B)和糖尿病性GK大鼠用SFN(GKS,C)、PM(GKP,D)和SFN+PM(GKSP,E)处理后的Nrf2染色。面板(F)包含不同动脉组中绿色荧光的量化。(G)根据DAPI染色(蓝色)的定义,定位于核区的Nrf2标签(绿色)的荧光强度被量化,并表示为从多个肠系膜动脉段汇集的Wistar大鼠的对照平均值的百分比。(H)不同动脉组主动脉中总Nrf2(左面板)和核Nrf2蛋白水平的Western blot条带。主动脉中总Nrf2水平(I)和核Nrf2水平(J)的平均密度测量数据,标准化为各自的负荷控制。数据平均值为±SE。*P<0.05,**P<0.01,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组;#P<0.05,###P<0.001 vs GKP组。

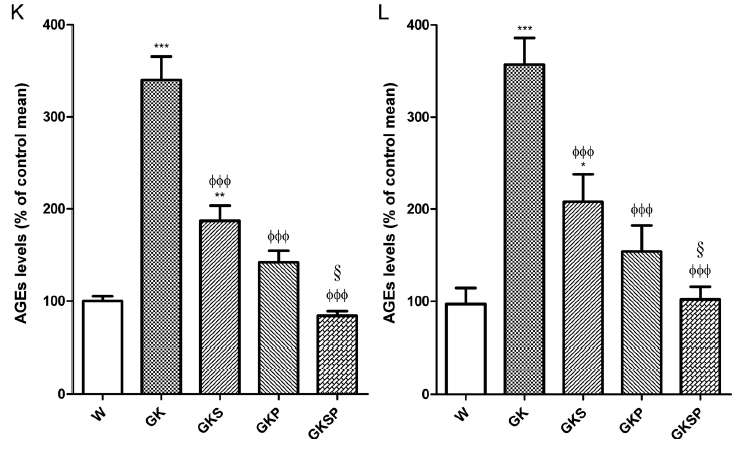 图7 与非糖尿病Wistar大鼠相比,萝卜硫素和吡哆胺对GK大鼠主动脉和肠系膜动脉AGE水平的影响。有代表性的主动脉切片显示非糖尿病Wistar大鼠(A)、糖尿病GK大鼠(B)和糖尿病GK大鼠经SFN(GKS,C)、PM(GKP,D)和SFN+PM(GKSP,E)处理后的AGE染色。面板(K)包含不同动脉组中红色荧光的量化。有代表性的肠系膜切片显示了非糖尿病Wistar大鼠(F)、糖尿病GK大鼠(G)和糖尿病GK大鼠用SFN(GKS,H)、PM(GKP,I)和SFN+PM(GKSP,J)处理后的AGE染色。面板(L)包含不同动脉组中红色荧光的量化。数据平均值为±SE。*P<0.05,**P<0.01,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组;§p<0.05 vs GKS组。
图7 与非糖尿病Wistar大鼠相比,萝卜硫素和吡哆胺对GK大鼠主动脉和肠系膜动脉AGE水平的影响。有代表性的主动脉切片显示非糖尿病Wistar大鼠(A)、糖尿病GK大鼠(B)和糖尿病GK大鼠经SFN(GKS,C)、PM(GKP,D)和SFN+PM(GKSP,E)处理后的AGE染色。面板(K)包含不同动脉组中红色荧光的量化。有代表性的肠系膜切片显示了非糖尿病Wistar大鼠(F)、糖尿病GK大鼠(G)和糖尿病GK大鼠用SFN(GKS,H)、PM(GKP,I)和SFN+PM(GKSP,J)处理后的AGE染色。面板(L)包含不同动脉组中红色荧光的量化。数据平均值为±SE。*P<0.05,**P<0.01,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组;§p<0.05 vs GKS组。
在糖尿病动物中,氧化应激与AGE的累积成比例增加,其导致血管并发症的发展。PM已被证明能抑制AGE的形成和脂源性美拉德产物(一种ALE)的形成。它不会直接与葡糖胺重排反应的产物相互作用,但会干扰重排后的氧化反应。此外,PM还可以捕获活性羰基化合物,抑制AGE和ALE加合物。PM抑制肥胖大鼠的脂血症,肾与血管并发症的发生。PM降低氧化应激的部分原因可能是抑制糖基化作用,而糖基化作用是一种直接导致自由基产生的过程。
此外,使用SFN、PM或两者治疗均能显著降低血管氧化应激,同时增加NO生物利用度,这可以部分解释对血管功能的有益影响。AGE直接阻断血管内皮的NO活性并产生活性氧。糖尿病环境的特点是血管的NO水平较低,其应归于氧化应激和AGE水平增加。SFN和PM对于降低氧化应激,正常化NO生物利用度非常有效。可能只有当AGE与ALE水平以及交联形成被PM降低,同时抗氧化防御酶(SFN作用)增加时,才有可能促成血管系统中更高水平的NO。
SFN对依赖Nrf2基因的表达的影响已被充分记录。重要的是,GK大鼠在血管水平上显示出Nrf2的缺失。类似地,在小鼠糖尿病动脉和人心脏组织中观察到Nrf2水平降低。此外,先前的研究表明,衰老与血管系统中的Nrf2功能障碍有关,这可能加剧与年龄相关的细胞氧化应激,并增加老化血管对氧化应激诱导的细胞损伤的敏感性。与这些发现一致,我们还发现8个月大的糖尿病GK大鼠主动脉和肠系膜动脉中的Nrf2表达显著下调,同时细胞核功能显著下降。Nrf2减少的机制尚不清楚,其解释也超出了当前研究的范围;然而,我们的数据表明,在糖尿病的血管系统中发生Nrf2信号传导的中断,它可能导致谷胱甘肽的消耗和其他抗氧化防御机制的减少。此外,以前的研究表明,Nrf2诱导了乙二醛酶1的表达,该酶可保护机体免受AGE诱导的蛋白质和DNA损伤,并维持细胞功能。
利用与抑制AGE/ALE水平和交联形成相关的Nrf2激动剂,并描述其有益作用,可为糖尿病及其血管并发症中恢复细胞内稳态提供一种新的治疗策略。据推测,这些发现导致了新的抗氧化分子的发现和评估,例如Nrf2激动剂和AGE抑制剂,它们可望在早期抑制糖尿病并发症的发病机制。
结论
总之,我们的研究结果表明,SFN+PM治疗内皮功能障碍的益处是多因素的。除了抗氧化功能降低氧化应激外,Nrf2还积极调节NO生物利用度。这些结果提供了令人信服的实验证据,证明SFN可与PM联用或不联用,改善内皮功能障碍和减轻糖尿病引起的血管损伤。在患有长期糖尿病的动物模型中,随着Nrf2的激活,糖基化降低,内皮功能正常化,这突出了靶向不同机制以实现主要有益结果的重要性。
综上所述,我们可以设想未来的策略,不仅包括早期积极治疗高血糖症,还包括同时使用针对AGE形成的活性化合物,以及能够特异性地针对反应性物种或增强我们抗氧化防御系统的化合物。同时激活Nrf2和抑制AGE/ALE水平与交联形成可能是一种潜在治疗策略。
方法
药物。PM、苯肾上腺素、乙酰胆碱、N-硝基-L-精氨酸甲酯(L-NAME)和超氧化物歧化酶-聚乙烯乙二醇购于SIGMA(St. Louis, MO, USA)。血管舒张剂激活磷蛋白(VASP)、磷酸化血管舒张剂激活磷蛋白(pVASP),β-肌动蛋白、Nrf2、AGE(克隆,No. 6D12)和硝基酪氨酸分别购于Cell Signaling Technology (Danvers, MA, USA)、Abcam plc(Cambridge, UK)、Santa Cruz Biotechnology(Santa Cruz, CA, USA)、Upstate Biotechnology(Lake placid, NY, USA)与Transgenic Inc(Kobe, Japan)。萝卜硫素(SFN,L异构体)购于LKT laboratories(St. Paul, MN),DHE购于Invitrogen(Barcelona, Spain)。本研究所用的所有其他化合物与试剂为优级纯。
实验动物。Wistar和GK大鼠来自我们当地的繁殖群体(葡萄牙科英布拉大学医学院)。将Wistar和GK糖尿病大鼠分为四个实验组。(1)组(W;GK)用载体(玉米油)维持8周;(2)组用萝卜硫素(1 mg/kg/天,腹腔内)治疗8周(WS;GKS);(3)组用吡哆胺(100 mg/kg/天,如前)治疗8周(WP;GKP);(4)组用萝卜硫素和吡哆胺治疗8周(WSP;GKSP)。这些动物用一种标准的商业颗粒饲料进行饲养,8个月大时使用。一项初步研究(数据未显示)表明,1 mg/kg在代谢和内皮功能方面具有更好的效果,并且接近通过食用西兰花芽苗提取物在人和小鼠体内获得的生理浓度。
所有动物均按照葡萄牙《实验动物实验法》(2004年最后修正案)接受护理,该法基于欧盟指令2010/63/EU对动物实验采用的实验动物护理原则。实验方案由科英布拉大学医学院ORBEA – IBILI批准。
代谢和氧化应激参数的测定。禁食15小时后,用氯胺酮/氯丙嗪[氯胺酮氯化物(75 mg/kg,im,Parke Davis,Ann Arbor,mi,USA)和氯丙嗪氯化物(2.65 mg/kg,im,lab.vit_ria,Portugal)]麻醉动物并斩首致死。心脏穿刺取血测定血脂。空腹血清脂质(总胆固醇和高密度脂蛋白胆固醇和甘油三酯)使用商业可获得的试剂盒下雨(奥林巴斯-塔格恩·斯塔卡葡萄牙,Olympus-Diagnóstica Portugal, Produtos de Diagnóstico SA, Portugal,葡萄牙)进行定量。血浆游离脂肪酸(FFA)水平使用酶分析试剂盒(Roche Applied Science, Portugal)进行评估。将大鼠置于代谢笼中24小时并收集尿液。尿8-羟基-2-脱氧鸟苷使用EIA试剂盒(Cayman Chemical, Europe)测量。
对于葡萄糖耐量试验,大鼠先前禁食过夜,并给予腹腔注射葡萄糖(1.75 g kg-1体重)。在注射葡萄糖后0、30、60和120分钟,通过葡萄糖氧化酶法从尾静脉中取样,使用葡萄糖仪(Glucometer Elite Bayer,Portugal S.A.)和兼容的反应测试条测定血糖。用标准计算机程序(GRAPPAD PRISM PCSoftware version 3.0)计算IPGTT曲线的AUC。糖化血红蛋白(HbA1c)使用A1C NOW+系统测定(Bayer,Portugal S.A.)。
等长张力研究。主动脉被迅速切除,去除结缔组织。主动脉分为两段(4-mm宽)。将环段安装在不锈钢三角板之间的单个器官室中,填充含氧(95%氧气,5%二氧化碳)改良Krebs-Henseleit缓冲液(37°C,pH7.4)(成分单位:mM:NaCl 119;KCl 4.7;CaCl2 1.6;MgSO4 1.2;NaHCO3 25;KH2PO4 1.2;葡萄糖 11.0)。在抑制前列腺素合成的实验中,吲哚美辛的浓度为10μM。主动脉环的静息张力为9.8 mN。平衡60分钟后,所有血管均预先用苯肾上腺素进行收缩。配体刺激受体介导的NO生物利用度通过对乙酰胆碱(Ach,10-9到10-3 M)的浓度依赖性松弛来评估,而硝普钠(SNP,10-9到10-4 M)被用作非内皮依赖性激动剂。对ACh和SNP的舒张反应以亚最大苯肾上腺素诱导收缩的舒张百分比表示,如前所述获得量效曲线。
根据Mulvani和Halpern的技术,切除肠系膜上二级动脉段(约200μm的外径),使其不含脂肪和粘附结缔组织,并安装在肌电图上。等长张力由PowerLab数据采集系统(Adinstruments,UK)记录和采集,并使用Labchart 7数据采集和分析软件(Adinstruments,UK)记录在计算机上。动脉段在改良的Krebs-Henseleit溶液中平衡30分钟,保持在37°C,并用5%的二氧化碳进行充气。根据Mulvani和Halpern的技术,将每只血管放置在相当于每只个体大鼠体内动脉血压的拉伸下。通过暴露于KCl(125 mM)评估组织收缩性;随后获得苯肾上腺素(0.1-30 μM)的累积量效曲线。让组织重新平衡30分钟。然后使用肠系膜环研究对以下一种药物的反应:对乙酰胆碱(Ach,10-9到10-6 M)的反应(在存在和不存在吲哚美辛(10 μM))或硝普钠(SNP,10-9到10-3 M)评估舒张;舒张根据先前的苯肾上腺素反应曲线,在使用苯肾上腺素预施工后,反应达到最大值的50%左右。在使用肠系膜动脉段的第二系列实验中,使用L-NAME评估了一氧化氮合酶抑制的急性效应。如上文所述获得对乙酰胆碱的反应,随后在L-NAME(300 μM)存在下重复。对ACh和SNP的舒张反应以亚最大苯肾上腺素诱导收缩的舒张百分比表示,并获得如前所述的浓度-响应曲线。
超氧化物检测。在37°C的PBS中用DHE(2×10−6 M)在避光加湿室中培养30分钟未固定的、30 μm厚的近端主动脉段。DHE在与氧反应后被氧化成溴化乙锭,溴化乙锭与细胞核中的DNA结合,发出红色荧光。对于溴化乙锭的检测,使用荧光显微镜(Leica Dmire200,Wetzlar,德国)获得图像。用568 nm滤光片检测荧光。正常组织和糖尿病组织的处理和成像使用相同的设置平行进行。所有制剂的显微镜和照相机设置均保持不变。荧光定量使用IMAGEJ(1.40 g,NIH)。
肠系膜免疫荧光的评估。将肠系膜动脉切片(6μm)用PBS洗涤,固定在冰冻丙酮中10分钟。随后在1% Triton X-100且pH值7.4的PBS中透化,用10%羊血清封闭10分钟,30分钟后,在含有0.02% BSA的PBS中稀释一次抗体(PBS/BSA)。添加一级抗体,并在4°C下将切片孵育过夜。孵育后,用PBS/BSA溶液对切片进行广泛清洗。之后,用二级抗体孵育切片,在PBS/BSA中稀释1小时。再用甘油凝胶Dako安装介质(Dako, Carpinteria, CA, USA),安装之前清洗盖片。免疫染色肠系膜切片用Leica-Dmire200荧光显微镜观察。免疫染色肠系膜切片用DAPI复染,并按上述方法检查、拍照和定量DHE荧光。
Western blot分析。用冷PBS清洗内皮完整血管段,并在缓冲液中冷却,缓冲液中含有:Tris-HCl 50、Nacl 150、乙二胺四乙酸(EDTA)1、乙二醇四乙酸(EGTA)0.1、NP-40、0.1%、SDS 0.1%和脱氧胆酸0.5%。苯甲基磺酰氟(1 mM)、抑肽酶(10μg ml-1)、亮蛋白(10μg ml-1)和胃蛋白酶抑制剂(10μg ml-1)均购于Sigma Chemicals(St. Louis, MO, USA),作为蛋白酶抑制剂加入。以标准方式对组织进行匀浆,然后在14000×g、4°C条件下离心20分钟。收集上清液并测定总蛋白浓度。将含有40μg蛋白质的样品加载到12%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)凝胶上,在聚偏氟乙烯膜上运行并电沉积。预染分子量标记蛋白被用作SDS-PAGE的标准。进行丽春红染色,以验证转移的质量,确保蛋白质负载相同。印迹用5%脱脂牛乳在PBS中封闭1h,用抗VASP、pVASP、β-肌动蛋白或Nrf2的一级抗体处理过夜,再用碱性磷酸酶二级抗体孵育1h,用抗VASP磷酸丝氨酸239抗体分析VASP在Ser239(pVASP)的磷酸化状态,这是一种血管cGMP依赖性蛋白激酶-1活性的可靠生化标志物。血管紧张素的激活由强度比pVASP/tVASP表示。免疫印迹是用ECF免疫印迹检测系统(Amersham Biosciences)开发的。
使用Bio-Rad蛋白质分析试剂盒测定蛋白质含量。
统计分析。所有数据均采用标准计算机程序(graphpad prism PC软件版本3.0,ANOVA)进行分析,并以平均值±SEM表示(n=12,每组个体动物数)。采用单因素方差分析,随后进行个体比较的Bonferroni事后检验,评估显著性差异。P<0.05被认为是显著的。用单纯形算法对剂量响应曲线进行非线性回归拟合。舒张反应以苯肾上腺素预收缩的百分比表示。量效曲线的比较采用双向方差分析对重复测量进行评估,随后进行个体比较的Bonferroni事后检验。
在本研究中,我们把吡哆胺(PM)和/或萝卜硫素(SFN)作为治疗干预措施,用于确定NFE2相关因子2(Nrf2)的激动剂是否可以同晚期糖基化终产物(AGE)形成抑制剂一起使用,达到减轻2型糖尿病患者的氧化应激和改善其内皮功能障碍。Goto Kakizaki(GK)大鼠,一种非肥胖型2型糖尿病动物模型,在8周内接受或不接受PM和/或SFN治疗,并与年龄匹配的Wistar大鼠进行比较。在治疗结束时,评估孤立主动脉和肠系膜动脉中一氧化氮(NO)依赖性和非依赖性的血管舒张。还评估了代谢概况、NO生物利用度和血管氧化应激、AGE和Nrf2水平。糖尿病GK大鼠的Nrf2水平显著降低,同时氧化应激和内皮功能障碍水平也较高。PM和SFN作为单一疗法能够显著改善主动脉和肠系膜动脉的内皮功能障碍,降低血管氧化损伤、AGE和HbA1c水平。此外,在GK大鼠中已证明,SFN+PM能更有效地降低全身游离脂肪酸水平,使内皮功能、NO生物利用度和糖化作用正常化。Nrf2激动剂可与AGE抑制剂和交联形成抑制剂联用来治疗2型糖尿病内皮功能障碍。
2型糖尿病与氧化应激水平升高和糖基化改变血管功能有关,因此被认为是心血管疾病发展的主要危险因素。许多的糖尿病并发症都与高血糖和活性氧物种(ROS)生成增加有关,其导致了内皮功能紊乱。对血管舒张剂(如乙酰胆碱)的内皮依赖性舒张功能受损是1型和2型糖尿病实验模型中导管和阻力动脉的共同特征。我们先前发现糖尿病GK大鼠的氧化应激和糖基化增加,而这种变化导向内皮功能障碍。
糖尿病的治疗进展并不能有效地减少糖尿病相关的血管并发症。考虑到ROS在内皮功能中的关键作用,人们已经作出了相当大的努力来探索降低血管系统中ROS的治疗方法。核因子E2(红系衍生2)相关因子-2(Nrf2)是一种转录因子,在细胞对氧化应激的保护中起着关键作用。Nrf2被称为抗氧化剂反应的“主调节器”。通过Nrf2/抗氧化反应元件途径激动剂诱导内源性抗氧化酶可能是获得足够水平的抗氧化剂和减少氧化应激的一种有趣方法。膳食中的NRF2激动剂,如十字花科蔬菜中发现的异硫氰酸盐萝卜硫素(SFN),可以增加抗氧化防御,降低血压,抑制肾脏内促炎信号通路,并预防高血糖引起的内皮细胞代谢紊乱。在糖尿病模型中,Nrf2的激活降低了野生型小鼠的血糖水平,减少了糖尿病相关肾病,但在Nrf2缺乏的小鼠中则没有。因此,Nrf2诱导剂在血管疾病中的治疗潜力巨大,特别是与长期2型糖尿病有关的血管疾病。
表1 8个月大的非糖尿病Wistar大鼠(W)、糖尿病Goto-Kakizaki大鼠(GK)、接受萝卜硫素处理的GK大鼠(GKS)、接受吡哆胺处理的GK大鼠(GKP)和接受萝卜硫素与吡哆胺处理的GK大鼠(GKSP)的体重和血脂水平。数据表示为平均值±SE(n=12每组动物)。*P<0.05,**P<0.01,***P<0.001 vs W大鼠。与GK大鼠相比,φφφP<0.001。§P<0.05 vs GKS大鼠。#P<0.05 vs GKP大鼠。
此外,在糖尿病情况下的晚期糖基化加速导致血管并发症。几位作者已经证明,抑制晚期糖基化终产物(AGE)形成可能是改善糖尿病血管并发症的一种治疗策略。吡哆胺(PM)是维生素B6的一种天然衍生物,已被证明是一种有效的蛋白质糖化和脂质氧化抑制剂。在体外和体内,PM均能抑制AGE的形成,对糖尿病肾病和视网膜病变的治疗具有潜在的益处。
萝卜硫素和吡哆胺有不同的作用机制。因此,我们假设单独或联合使用SFN、PM对2型糖尿病模型Goto-Kakizaki(GK)大鼠的内皮依赖性血管反应性、氧化应激和代谢参数具有有益影响。在糖尿病进展的同时,对脂质分布、氧化应激、糖基化和一氧化氮(NO)生物利用度进行了评价。此外,还评估了主动脉和肠系膜动脉对乙酰胆碱(ACH)和硝普钠(SNP)的内皮依赖性和非依赖性血管敏感性。与Wistar大鼠相比,8个月大的GK大鼠表现出长期糖尿病表型,导管和阻力动脉内皮功能障碍,以及氧化应激增加。单用SFN和PM可以改善孤立动脉中的NO依赖性血管舒张。顺带一提,SFN和PM通过一种机制使主动脉和肠系膜动脉的内皮功能正常化,该机制包括NO生物利用度的增加,以及氧化应激和AGE水平的降低。总之,这些研究表明,针对引起内皮功能障碍的不同机制,抑制AGE形成(PM)和促进抗氧化防御系统(SFN)增强应是对抗糖尿病大血管并发症的有用工具,因其同时作用于最终导致血管损伤的不同机制。
结论
动物特征。我们实验中使用的糖尿病大鼠在研究开始时表现出类似的空腹血糖水平。治疗前的GK大鼠血糖(GK6M)明显高于年龄匹配的非糖尿病大鼠(图1补充)。GK大鼠的体重明显低于年龄匹配的Wistar大鼠,不同的治疗方法并没有显著改变这个参数(表1)。在腹膜内葡萄糖耐量试验(IPGTT)中,GK大鼠与Wistar大鼠相比表现出明显的葡萄糖不耐(图1A)。与Wistar大鼠相比,GK大鼠的空腹血糖、葡萄糖曲线下面积(AUC)、HbA1c、总胆固醇和游离脂肪酸(FFA)升高(图1B、C、D、表1)。如前所述,GK大鼠表现出正常的非HDL胆固醇和甘油三酯水平(表1)。单用SFN或PM治疗并没有显著改变糖耐量和脂质分布评估(图1A、C,表1)。同时,PM显著降低了空腹血糖和HbA1c水平(图1B、D)。事实上,所有的治疗方法都能有效降低GK大鼠的HbA1c水平(图1D)。此外,使用SFN和PM治疗8周可有效降低糖尿病大鼠的血液中FFA浓度(表1),并显著改善葡萄糖耐受性(图1A、C)。
NO依赖性血管舒张。8个月大的GK大鼠与年龄匹配的Wistar大鼠相比,苯肾上腺素预收缩后的主动脉环响应乙酰胆碱(ACh)引起的内皮介导血管舒张能力受损(图2A),但对硝普钠(SNP)引起的非内皮依赖性舒张在这两个物种中相似(图3A)。事实上,在糖尿病大鼠中,苯肾上腺素预收缩主动脉环对乙酰胆碱引起的内皮介导血管最大舒张值降低了49%(图2A)。在GK大鼠和Wistar大鼠中,用NOS抑制剂N-硝基-L-精氨酸甲酯(L-NAME)和环氧合酶抑制剂吲哚美辛预孵化后的动脉环几乎完全消除了Ach的松弛作用(图2补充)。在我们的实验条件下,其他血管扩张剂的残余成分约为15%。在主动脉中,没有观察到不同组间SNP量效曲线在最大舒张值上的差异(图3A)。所有GK治疗组的最大舒张值显著改善,对Ach的血管反应性有所改善(图2A,表2)。用SFN、PM或SFN+PM治疗可显著改善糖尿病大鼠主动脉内皮依赖性血管舒张(图2A,表2)。这种血管舒张对吲哚美辛不敏感,并被L-NAME阻断,表明其机制依赖于内源性NO释放(图2补充)。此外,在所有GK治疗组中观察到SNP敏感性增加(图3A,表2)。在肠系膜动脉中,糖尿病大鼠的苯肾上腺素预收缩后动脉环响应Ach引起的内皮介导的最大舒张值降低了36%(图2B)。此外,糖尿病大鼠响应SNP引起的最大舒张值下降了39%(图3B)。用SFN、PM或SFN+PM治疗能够改善内皮依赖性血管扩张(图2B,表3)。类似地,这种作用对吲哚美辛不敏感,并被L-NAME抑制(图2补充)。表2和表3总结了最大舒张值和EC50值的详细数据。这些结果表明,单用SFN或PM治疗可改善糖尿病GK大鼠主动脉和肠系膜动脉的内皮依赖性血管舒张。与此相关,这些疗法能够使内皮介导的舒张正常化,并提高主动脉和肠系膜动脉的SNP敏感性(图2、3)。
图1 与非糖尿病Wistar(W)大鼠相比,8个月大的糖尿病Goto-Kakizaki(GK)大鼠萝卜硫素和吡哆胺处理的效果,腹腔葡萄糖耐量试验(IPGTT;A)、空腹血糖(B)、葡萄糖曲线下面积(AUC;C)和糖化血红蛋白(HbA1c)水平(D)。C)计算IPGTT曲线的AUC,以测量葡萄糖耐量受损的程度。数据表示为平均值±SE(n=12)。***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组。
血管壁的氧化应激。我们确定了SFN、PM或两者对糖尿病血管系统氧化应激的潜在影响。有趣的是,糖尿病导致糖尿病大动脉中的超氧化物生成量增加了3倍(p<0.001;图4B,K)。与GK大鼠相比,用SFN、PM和SFN+PM治疗的糖尿病大鼠主动脉中二氢乙啶(DHE,ROS荧光探针)的密度显著降低(图4C、D、E、K)。此外,糖尿病GK大鼠肠系膜动脉中的免疫反应性硝基酪氨酸(蛋白质氧化生物标志物)水平也升高(p<0.001;图4G,L),并且用SFN、PM或两者治疗后,这些水平显著降低(图4H,I,J,L)。因此,所有疗法都能显著降低糖尿病GK大鼠主动脉和肠系膜动脉的血管氧化损伤。相应地,GK大鼠尿中8-羟基-2’-脱氧鸟苷(8-OHdG,DNA氧化损伤生物标志物)的水平明显高于年龄匹配的Wistar大鼠(图4M)。对GK大鼠使用SFN、PM或两者治疗8周后,8-OHdG水平显著降低(图4M)。
主动脉中的肺血管紧张素和总血管紧张素。血管舒张剂激活磷蛋白(VASP)在Ser239的磷酸化被证明是生物活性NO和NO-cGMP蛋白激酶G信号通路活性的指标。为了确定NO生物利用度,我们测量了主动脉中磷酸化血管舒张剂激活磷蛋白(pVASP)和总VASP。GK大鼠的主动脉pVASP/tVASP比率明显低于Wistar大鼠(图5A,B)。在糖尿病GK大鼠(GKS,GKP;GKSP;图5A,B)中,用SFN、PM或两者治疗可显著提高pVASP/TVSP比率。在肠系膜动脉中观察到类似的情况(数据未列出)。
主动脉和肠系膜动脉中的Nrf2水平。我们随后量化了不同组大鼠动脉中的Nrf2水平。用Western blot检测主动脉中的Nrf2水平,以共聚焦显微镜观察肠系膜动脉中的免疫荧光检来测Nrf2水平。糖尿病大鼠血管中的Nrf2水平明显低于主动脉和肠系膜动脉中年龄匹配的Wistar大鼠(图6I,F)。萝卜硫素治疗组(GKS和GKSP)显著增加了主动脉和肠系膜动脉中的细胞核Nrf2水平(图6J,G)。Nrf2激动剂SFN增加糖尿病GK大鼠Nrf2的核定位,这可能导致抗氧化防御增加,促进血管氧化应激的减少。在糖尿病GK大鼠的主动脉和肠系膜动脉中,SFN显著增加了Nrf2的表达(图6),这表明它同时促进了表达增加和核易位,与其在其他组织中的作用相一致。
主动脉和肠系膜动脉的AGE水平。接下来,我们量化了不同组大鼠动脉中的AGE水平。主动脉和肠系膜动脉的AGE水平用共聚焦显微镜测量。糖尿病大鼠主动脉和肠系膜动脉中的AGE水平明显高于年龄匹配的Wistar大鼠(分别为图7B、K、G、L)。GK治疗组(GKS、GKP和GKSP)主动脉和肠系膜动脉的血管AGE水平显著降低(图7K,L)。值得注意的是,PM治疗组(GKP,GKSP)血管壁具有正常的AGE水平。
讨论
在目前的研究中,萝卜硫素、吡哆胺及其联用对GK大鼠的糖尿病内皮功能障碍表现出明显的治疗潜力。用SFN、PM或两者联合治疗GK大鼠可显著降低主动脉和肠系膜动脉的内皮功能障碍。在这些血管中,SFN、PM或SFN+PM治疗可延缓包括氧化损伤在内的糖尿病内皮功能障碍病理特征的出现。此外,这两种相关疗法都能降低FFA水平,也增加了NO生物利用度,这可以部分解释2型糖尿病长期模型中内皮功能的正常化。
GK大鼠为非肥胖2型糖尿病动物模型,具有轻度高血糖、高胰岛素血症和胰岛素抵抗。长期糖尿病通常与这些动物的胰腺功能丧失有关,其特征也类似于人类疾病进展中的相关特征。该模型与人类2型糖尿病共有包括异常血管反应性在内的多种心血管表型,证明其作为研究糖尿病并发症的模型是合适的。
在此,我们发现8个月大的GK大鼠具有高血糖,显著增加的总胆固醇和循环FFA水平的特征。内皮功能障碍在肠系膜动脉和主动脉动脉中都很明显,其伴随着氧化应激和糖基化作用升高、NO生物利用度下降和Nrf2水平降低。
Nrf2抗氧化功能在血管疾病中可能很重要。SFN是一种在西兰花中发现的Nrf2激动剂,增加抗氧化反应元件相关转录控制下的保护酶的表达,并防止由高血糖引起的内皮细胞代谢功能障碍。在其它研究的基础上,结合西兰花芽苗中SFN的生物利用度,测定了该化合物的浓度。我们估计,在人体饮食可行的生理剂量下,SFN对治疗内皮功能障碍具有潜在的治疗益处。另一方面,我们最近检查了2型糖尿病动物模型的AGE水平,发现血浆和血管壁的AGE水平都在增加。如PM的化合物可以降低高水平的AGE,和这种方法对内皮功能有潜在的有益作用。这两种疗法对大血管疾病的治疗效果尚未被研究过。在本文中,我们研究了在血管水平上,有或没有与PM共同抑制年龄形成的SFN的作用。
图4 与非糖尿病Wistar(W)大鼠相比,萝卜硫素和吡哆胺处理对GK大鼠全身和血管氧化应激的影响。代表性的DHE染色的主动脉段反映了不同处理(A-E)下的氧气生成。所有的内皮都朝上。在相同的条件下,糖尿病患者GK(B)的荧光明显高于正常血管(Wistar,A)。注意到GK主动脉内皮、内膜和介质中反射O2·−水平的荧光增强。糖尿病GK大鼠服用SFN(GKS,C)、PM(GKP,D)和SFN+PM(GKSP,E)后,DHE荧光降低。面板(K)包含不同动脉组中荧光溴化乙锭信号的量化。具有代表性的肠系膜切片用硝基酪氨酸染色显示非糖尿病Wistar大鼠(F),糖尿病GK大鼠(G),糖尿病GK大鼠SFN(GKS,H)、PM(GKP,I)和SFN+PM(GKSP,J)处理组。面板(L)包含不同动脉组中绿色荧光的量化。(M)不同组大鼠尿8-羟基脱氧鸟苷(8-OHDG)水平。数据表示为平均值±SE(n=12)。*P<0.05,***P<0.001 vs Wistar组;φP<0.05,φφP<0.01,φφφP<0.001 vs GK组。
单用SFN或PM治疗8周并没有显著改善甘油三酯、胆固醇和FFA水平。已经证明,PM可以纠正糖尿病和肥胖大鼠的高脂血症和肾病,可能是通过干扰各种AGE/晚期脂质氧化最终产物(ALE,由脂质氧化形成)的活性羰基中间产物。在不同的动物模型中,更高浓度的PM(400 mg/kg/天)具有这种降脂作用。此外,Nrf2通路具有调节肝脏脂质代谢的功能。Nrf2负性调节许多编码参与脂质生物合成、脂肪酸去饱和与脂肪酸转运的酶的基因。相反,0.5 mg/kg的SFN会损害链脲佐菌素诱导的糖尿病大鼠的肝功能并加重的胆固醇水平。此外,在我们的实验条件下,单用SFN治疗不会改变脂质分布。
SFN+PM治疗对总胆固醇和甘油三酯水平没有影响,但却显著降低了FFA和HbA1c水平。两种联合治疗均能改善葡萄糖耐受性,显著减少葡萄糖曲线下面积。靶向糖基化和抗氧化防御系统的PM和SFN在降低FFA和HbA1c水平方面非常有效。重要的是,SFN和PM治疗对胰岛素水平没有影响,这解释了在治疗糖尿病大鼠中观察到的高血糖。事实上,在所有GK治疗组中,血糖水平明显高于Wistar大鼠。然而,在这些条件下,sfn+pm的血管保护作用是明显的,表明正常血糖的完全恢复不是血管保护的先决条件。
其他研究者先前报道,在非缺陷Nrf2的野生型小鼠中,Nrf2的激活降低了血糖水平,减少了糖尿病相关肾病。而这项研究中,SFN治疗组(GKS)没有促进血糖水平下降。不同的动物模型和实验条件可以解释这些差异。
氧化应激在内皮功能障碍发病机制中的作用已被充分确证。糖尿病增加血管系统中的活性氧,损害抗氧化防御酶,并降低细胞内抗氧化物的水平,创造一个氧化应激增加的环境。在血管模型中,Nrf2通路激动剂通过增加抗氧化/亲电反应元件介导的II相酶和抗氧化酶表达(包括NAD(P)H:醌氧化还原酶-1、血红素氧合酶-1和γ-谷氨酸-半胱氨酸连接酶催化亚单位)来恢复氧化还原平衡。在这项研究中,我们提出的证据表明,SFN和PM治疗在体内具有抗氧化作用:在糖尿病GK大鼠中,SFN、PM或SFN+PM降低了尿8-OHdG和组织中O2•−阴离子以及硝基酪氨酸的积累量。这种抗氧化作用是已观察到的内皮功能障碍正常化机制的重要因素。因此,其他研究也报道了不同条件下SFN对氧化应激的抑制作用。在糖尿病db/db微血管系统中,用SFN靶向Nrf2通路能有效恢复正常的氧化还原平衡和对抵抗小动脉的肌源性反应。此外,通过介导上调Nrf2活性,SFN下调高脂饮食喂养的小鼠主动脉中血管细胞粘附分子-1的表达。我们的数据支持一个模型,在该模型中,通过一种涉及Nrf2依赖性的抗氧化能力下降的机制,主动脉和肠系膜动脉中的氧化应激增加。我们在活体内证明,用SFN靶向Nrf2通路可有效恢复正常的氧化还原平衡,改善糖尿病血管系统中主动脉和肠系膜动脉对乙酰胆碱的内皮反应。这些有益的影响被PM增强,而PM是一种AGE/ALE和其交联形成的抑制剂。
在糖尿病动物中,氧化应激与AGE的累积成比例增加,其导致血管并发症的发展。PM已被证明能抑制AGE的形成和脂源性美拉德产物(一种ALE)的形成。它不会直接与葡糖胺重排反应的产物相互作用,但会干扰重排后的氧化反应。此外,PM还可以捕获活性羰基化合物,抑制AGE和ALE加合物。PM抑制肥胖大鼠的脂血症,肾与血管并发症的发生。PM降低氧化应激的部分原因可能是抑制糖基化作用,而糖基化作用是一种直接导致自由基产生的过程。
此外,使用SFN、PM或两者治疗均能显著降低血管氧化应激,同时增加NO生物利用度,这可以部分解释对血管功能的有益影响。AGE直接阻断血管内皮的NO活性并产生活性氧。糖尿病环境的特点是血管的NO水平较低,其应归于氧化应激和AGE水平增加。SFN和PM对于降低氧化应激,正常化NO生物利用度非常有效。可能只有当AGE与ALE水平以及交联形成被PM降低,同时抗氧化防御酶(SFN作用)增加时,才有可能促成血管系统中更高水平的NO。
SFN对依赖Nrf2基因的表达的影响已被充分记录。重要的是,GK大鼠在血管水平上显示出Nrf2的缺失。类似地,在小鼠糖尿病动脉和人心脏组织中观察到Nrf2水平降低。此外,先前的研究表明,衰老与血管系统中的Nrf2功能障碍有关,这可能加剧与年龄相关的细胞氧化应激,并增加老化血管对氧化应激诱导的细胞损伤的敏感性。与这些发现一致,我们还发现8个月大的糖尿病GK大鼠主动脉和肠系膜动脉中的Nrf2表达显著下调,同时细胞核功能显著下降。Nrf2减少的机制尚不清楚,其解释也超出了当前研究的范围;然而,我们的数据表明,在糖尿病的血管系统中发生Nrf2信号传导的中断,它可能导致谷胱甘肽的消耗和其他抗氧化防御机制的减少。此外,以前的研究表明,Nrf2诱导了乙二醛酶1的表达,该酶可保护机体免受AGE诱导的蛋白质和DNA损伤,并维持细胞功能。
利用与抑制AGE/ALE水平和交联形成相关的Nrf2激动剂,并描述其有益作用,可为糖尿病及其血管并发症中恢复细胞内稳态提供一种新的治疗策略。据推测,这些发现导致了新的抗氧化分子的发现和评估,例如Nrf2激动剂和AGE抑制剂,它们可望在早期抑制糖尿病并发症的发病机制。
结论
总之,我们的研究结果表明,SFN+PM治疗内皮功能障碍的益处是多因素的。除了抗氧化功能降低氧化应激外,Nrf2还积极调节NO生物利用度。这些结果提供了令人信服的实验证据,证明SFN可与PM联用或不联用,改善内皮功能障碍和减轻糖尿病引起的血管损伤。在患有长期糖尿病的动物模型中,随着Nrf2的激活,糖基化降低,内皮功能正常化,这突出了靶向不同机制以实现主要有益结果的重要性。
综上所述,我们可以设想未来的策略,不仅包括早期积极治疗高血糖症,还包括同时使用针对AGE形成的活性化合物,以及能够特异性地针对反应性物种或增强我们抗氧化防御系统的化合物。同时激活Nrf2和抑制AGE/ALE水平与交联形成可能是一种潜在治疗策略。
方法
药物。PM、苯肾上腺素、乙酰胆碱、N-硝基-L-精氨酸甲酯(L-NAME)和超氧化物歧化酶-聚乙烯乙二醇购于SIGMA(St. Louis, MO, USA)。血管舒张剂激活磷蛋白(VASP)、磷酸化血管舒张剂激活磷蛋白(pVASP),β-肌动蛋白、Nrf2、AGE(克隆,No. 6D12)和硝基酪氨酸分别购于Cell Signaling Technology (Danvers, MA, USA)、Abcam plc(Cambridge, UK)、Santa Cruz Biotechnology(Santa Cruz, CA, USA)、Upstate Biotechnology(Lake placid, NY, USA)与Transgenic Inc(Kobe, Japan)。萝卜硫素(SFN,L异构体)购于LKT laboratories(St. Paul, MN),DHE购于Invitrogen(Barcelona, Spain)。本研究所用的所有其他化合物与试剂为优级纯。
实验动物。Wistar和GK大鼠来自我们当地的繁殖群体(葡萄牙科英布拉大学医学院)。将Wistar和GK糖尿病大鼠分为四个实验组。(1)组(W;GK)用载体(玉米油)维持8周;(2)组用萝卜硫素(1 mg/kg/天,腹腔内)治疗8周(WS;GKS);(3)组用吡哆胺(100 mg/kg/天,如前)治疗8周(WP;GKP);(4)组用萝卜硫素和吡哆胺治疗8周(WSP;GKSP)。这些动物用一种标准的商业颗粒饲料进行饲养,8个月大时使用。一项初步研究(数据未显示)表明,1 mg/kg在代谢和内皮功能方面具有更好的效果,并且接近通过食用西兰花芽苗提取物在人和小鼠体内获得的生理浓度。
所有动物均按照葡萄牙《实验动物实验法》(2004年最后修正案)接受护理,该法基于欧盟指令2010/63/EU对动物实验采用的实验动物护理原则。实验方案由科英布拉大学医学院ORBEA – IBILI批准。
代谢和氧化应激参数的测定。禁食15小时后,用氯胺酮/氯丙嗪[氯胺酮氯化物(75 mg/kg,im,Parke Davis,Ann Arbor,mi,USA)和氯丙嗪氯化物(2.65 mg/kg,im,lab.vit_ria,Portugal)]麻醉动物并斩首致死。心脏穿刺取血测定血脂。空腹血清脂质(总胆固醇和高密度脂蛋白胆固醇和甘油三酯)使用商业可获得的试剂盒下雨(奥林巴斯-塔格恩·斯塔卡葡萄牙,Olympus-Diagnóstica Portugal, Produtos de Diagnóstico SA, Portugal,葡萄牙)进行定量。血浆游离脂肪酸(FFA)水平使用酶分析试剂盒(Roche Applied Science, Portugal)进行评估。将大鼠置于代谢笼中24小时并收集尿液。尿8-羟基-2-脱氧鸟苷使用EIA试剂盒(Cayman Chemical, Europe)测量。
对于葡萄糖耐量试验,大鼠先前禁食过夜,并给予腹腔注射葡萄糖(1.75 g kg-1体重)。在注射葡萄糖后0、30、60和120分钟,通过葡萄糖氧化酶法从尾静脉中取样,使用葡萄糖仪(Glucometer Elite Bayer,Portugal S.A.)和兼容的反应测试条测定血糖。用标准计算机程序(GRAPPAD PRISM PCSoftware version 3.0)计算IPGTT曲线的AUC。糖化血红蛋白(HbA1c)使用A1C NOW+系统测定(Bayer,Portugal S.A.)。
等长张力研究。主动脉被迅速切除,去除结缔组织。主动脉分为两段(4-mm宽)。将环段安装在不锈钢三角板之间的单个器官室中,填充含氧(95%氧气,5%二氧化碳)改良Krebs-Henseleit缓冲液(37°C,pH7.4)(成分单位:mM:NaCl 119;KCl 4.7;CaCl2 1.6;MgSO4 1.2;NaHCO3 25;KH2PO4 1.2;葡萄糖 11.0)。在抑制前列腺素合成的实验中,吲哚美辛的浓度为10μM。主动脉环的静息张力为9.8 mN。平衡60分钟后,所有血管均预先用苯肾上腺素进行收缩。配体刺激受体介导的NO生物利用度通过对乙酰胆碱(Ach,10-9到10-3 M)的浓度依赖性松弛来评估,而硝普钠(SNP,10-9到10-4 M)被用作非内皮依赖性激动剂。对ACh和SNP的舒张反应以亚最大苯肾上腺素诱导收缩的舒张百分比表示,如前所述获得量效曲线。
根据Mulvani和Halpern的技术,切除肠系膜上二级动脉段(约200μm的外径),使其不含脂肪和粘附结缔组织,并安装在肌电图上。等长张力由PowerLab数据采集系统(Adinstruments,UK)记录和采集,并使用Labchart 7数据采集和分析软件(Adinstruments,UK)记录在计算机上。动脉段在改良的Krebs-Henseleit溶液中平衡30分钟,保持在37°C,并用5%的二氧化碳进行充气。根据Mulvani和Halpern的技术,将每只血管放置在相当于每只个体大鼠体内动脉血压的拉伸下。通过暴露于KCl(125 mM)评估组织收缩性;随后获得苯肾上腺素(0.1-30 μM)的累积量效曲线。让组织重新平衡30分钟。然后使用肠系膜环研究对以下一种药物的反应:对乙酰胆碱(Ach,10-9到10-6 M)的反应(在存在和不存在吲哚美辛(10 μM))或硝普钠(SNP,10-9到10-3 M)评估舒张;舒张根据先前的苯肾上腺素反应曲线,在使用苯肾上腺素预施工后,反应达到最大值的50%左右。在使用肠系膜动脉段的第二系列实验中,使用L-NAME评估了一氧化氮合酶抑制的急性效应。如上文所述获得对乙酰胆碱的反应,随后在L-NAME(300 μM)存在下重复。对ACh和SNP的舒张反应以亚最大苯肾上腺素诱导收缩的舒张百分比表示,并获得如前所述的浓度-响应曲线。
超氧化物检测。在37°C的PBS中用DHE(2×10−6 M)在避光加湿室中培养30分钟未固定的、30 μm厚的近端主动脉段。DHE在与氧反应后被氧化成溴化乙锭,溴化乙锭与细胞核中的DNA结合,发出红色荧光。对于溴化乙锭的检测,使用荧光显微镜(Leica Dmire200,Wetzlar,德国)获得图像。用568 nm滤光片检测荧光。正常组织和糖尿病组织的处理和成像使用相同的设置平行进行。所有制剂的显微镜和照相机设置均保持不变。荧光定量使用IMAGEJ(1.40 g,NIH)。
肠系膜免疫荧光的评估。将肠系膜动脉切片(6μm)用PBS洗涤,固定在冰冻丙酮中10分钟。随后在1% Triton X-100且pH值7.4的PBS中透化,用10%羊血清封闭10分钟,30分钟后,在含有0.02% BSA的PBS中稀释一次抗体(PBS/BSA)。添加一级抗体,并在4°C下将切片孵育过夜。孵育后,用PBS/BSA溶液对切片进行广泛清洗。之后,用二级抗体孵育切片,在PBS/BSA中稀释1小时。再用甘油凝胶Dako安装介质(Dako, Carpinteria, CA, USA),安装之前清洗盖片。免疫染色肠系膜切片用Leica-Dmire200荧光显微镜观察。免疫染色肠系膜切片用DAPI复染,并按上述方法检查、拍照和定量DHE荧光。
Western blot分析。用冷PBS清洗内皮完整血管段,并在缓冲液中冷却,缓冲液中含有:Tris-HCl 50、Nacl 150、乙二胺四乙酸(EDTA)1、乙二醇四乙酸(EGTA)0.1、NP-40、0.1%、SDS 0.1%和脱氧胆酸0.5%。苯甲基磺酰氟(1 mM)、抑肽酶(10μg ml-1)、亮蛋白(10μg ml-1)和胃蛋白酶抑制剂(10μg ml-1)均购于Sigma Chemicals(St. Louis, MO, USA),作为蛋白酶抑制剂加入。以标准方式对组织进行匀浆,然后在14000×g、4°C条件下离心20分钟。收集上清液并测定总蛋白浓度。将含有40μg蛋白质的样品加载到12%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)凝胶上,在聚偏氟乙烯膜上运行并电沉积。预染分子量标记蛋白被用作SDS-PAGE的标准。进行丽春红染色,以验证转移的质量,确保蛋白质负载相同。印迹用5%脱脂牛乳在PBS中封闭1h,用抗VASP、pVASP、β-肌动蛋白或Nrf2的一级抗体处理过夜,再用碱性磷酸酶二级抗体孵育1h,用抗VASP磷酸丝氨酸239抗体分析VASP在Ser239(pVASP)的磷酸化状态,这是一种血管cGMP依赖性蛋白激酶-1活性的可靠生化标志物。血管紧张素的激活由强度比pVASP/tVASP表示。免疫印迹是用ECF免疫印迹检测系统(Amersham Biosciences)开发的。
使用Bio-Rad蛋白质分析试剂盒测定蛋白质含量。
统计分析。所有数据均采用标准计算机程序(graphpad prism PC软件版本3.0,ANOVA)进行分析,并以平均值±SEM表示(n=12,每组个体动物数)。采用单因素方差分析,随后进行个体比较的Bonferroni事后检验,评估显著性差异。P<0.05被认为是显著的。用单纯形算法对剂量响应曲线进行非线性回归拟合。舒张反应以苯肾上腺素预收缩的百分比表示。量效曲线的比较采用双向方差分析对重复测量进行评估,随后进行个体比较的Bonferroni事后检验。
本文由福山生物整理翻译,转载请注明出处。