






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
Keap1-Nrf2信号在情绪障碍中的重要作用:综述与展望
发表于:2020-04-28 作者:admin 来源:本站 点击量:14489
Essential Role of Keap1-Nrf2 Signaling in Mood Disorders: Overview and Future Perspective
(Keap1-Nrf2信号在情绪障碍中的重要作用:综述和未来展望)
(Keap1-Nrf2信号在情绪障碍中的重要作用:综述和未来展望)
摘要
抑郁症是最常见的心境障碍之一,复发率很高。积累的证据表明,Kelch样红系细胞CNC同源性衍生蛋白(ECH)相关蛋白1(Keap1)-核因子(红系衍生2)-样2(Nrf2)系统在与抑郁症相关的炎症中起着关键作用。临床前研究表明,抑郁样表型小鼠海马额叶前部皮层(PFC)、CA3区和齿状回(DG)中的Keap1和Nrf2蛋白表达低于对照小鼠。在习得性无助性范式中,抑郁样表型大鼠海马PFC和DG中的Keap1和Nrf2蛋白水平也低于对照大鼠和弹性大鼠。此外,具有抑郁样表型的啮齿动物促炎细胞因子水平更高。有趣的是,与野生型小鼠相比,Nrf2敲除(KO)小鼠表现出抑郁样表型和更高的血清促炎性细胞因子水平。同时,与野生型小鼠相比,Nrf2-KO小鼠的脑源性神经营养因子(BDNF)在海马PFC、CA3区和DG中的表达较低。7,8-二羟基黄酮是一种TrkB激动剂,刺激PFC、CA3区和DG中的BDNF-TrkB,在Nrf2-KO小鼠中显示出抗抑郁作用。用天然的Nrf2激动剂萝卜硫素进行预处理,可以防止炎症或慢性社会挫败应激后的小鼠出现抑郁样表型。有趣的是,在小鼠幼年期和青少年期饮食中摄入含0.1%萝卜硫苷(萝卜硫素的前体)的食物可以预防成年期小鼠慢性社会挫败应激后的抑郁样表型。此外,严重抑郁症和双相情感障碍患者的顶叶皮层中Keap1和Nrf2的蛋白表达低于对照组。这些发现表明,Keap1-Nrf2系统在应激弹性中起着关键作用,这与心境障碍的病理生理学有关。因此,饮食中摄入包含萝卜硫苷(或萝卜硫素) 十字花科蔬菜,可能预防或尽量减少抑郁症患者因应激和/或炎症引起的缓解后复发。在综述中,作者将讨论Keap1-Nrf2系统在情绪障碍中的作用。
简介
抑郁症是世界上最常见的精神疾病之一,是一种高复发率的心境障碍。世界卫生组织(WHO)估计,各年龄段共超过3.2亿人患有抑郁症,并强调这种疾病是全球疾病负担的主要原因(World Health Organization [WHO], 2017)。尽管目前还不清楚抑郁症病理生理学的确切机制,但积累的证据表明,抑郁症的病理生理学中存在炎症过程(Dantzer et al., 2008; Hashimoto, 2009; Miller et al., 2009; Raison et al., 2010; Hashimoto, 2015; Mechawar and Savitz, 2016; Miller and Raison, 2016; Zhang et al., 2016; Miller et al., 2017)。荟萃分析显示,与健康对照组相比,进行或不进行药物治疗的抑郁症患者血液中促炎细胞因子水平更高(Dowlati et al., 2010; Young et al., 2014; Haapakoski et al., 2015; Köhler et al., 2018)。研究表明,抑郁症患者死后的大脑样本中促炎细胞因子的基因表达水平升高(Dean et al., 2010; Shelton et al., 2011)。总之,炎症可能在抑郁症的病理生理学中起着关键作用。
在过去的十年里,人们对早期干预心境障碍的潜在益处越来越感兴趣。一些证据表明,营养对抑郁症的发展有很大影响(Lin et al., 2010; Murakami and Sasaki, 2010; Bazinet and Layé, 2014; Mello et al., 2014; El-Behadli et al., 2015; Opie et al., 2015; Lin et al., 2017; Hsu et al., 2018; Wang et al., 2018)。最近的荟萃分析表明,水果、蔬菜、鱼类和全谷物的高摄入量与较低的抑郁症风险有关(Lai et al., 2014; Liu et al., 2016; Saghafian et al., 2018)。
在本综述中,作者将讨论Keap1(Kelch样红系细胞CNC同源衍生蛋白[ECH]-相关蛋白1)]-Nrf2(核因子(红系衍生)样2)系统在抑郁症病理生理学中的作用,因为Keap1-Nrf2系统在炎症中起着关键作用。此外,我们还提到了天然Nrf2激动剂萝卜硫素(SFN)作为心境障碍营养干预的临床意义。
Keap1-Nrf2系统
Nrf2是一种转录因子,在细胞抵御氧化应激中起着关键作用。它与抗氧化反应元件(ARE)结合,ARE位于许多II相解毒或抗氧化酶及相关应激反应蛋白的编码基因的启动子区(Kobayashi et al., 2013; Ma, 2013; Suzuki et al., 2013a; Suzuki and Yamamoto, 2015; Yamamoto et al., 2018)。在正常条件下,Nrf2被Keap1抑制,Keap1是Nrf2降解的衔接蛋白(Suzuki等人,2013a;Suzuki和Yamamoto,2015)。在氧化应激期间,Nrf2去抑制并激活保护基因的转录(Suzuki et al., 2013a; Suzuki and Yamamoto, 2015)。重要的是,Keap1-Nrf系统在炎症相关的发病机制中起着关键作用(Kobayashi et al., 2013; Suzuki et al., 2013a; O’Connell and Hayes, 2015; Suzuki and Yamamoto, 2015; Wardyn et al., 2015; Yamamoto et al., 2018)。在癌细胞中,Nrf2激活对癌症承载宿主是有益的还是有害的,取决于时间(启动、促进和转移)和位置(癌细胞或微环境)(Yamamoto et al., 2018)。
Nrf2激动剂
基于Nrf2在预防各种病理状况中的作用,我们努力从天然来源中分离或开发有效且特异的Nrf2激动剂(Yamamoto et al., 2018)。天然强效抗炎化合物萝卜硫素(SFN:1-异硫氰酸-4-甲基亚磺酰丁烷)是一种有机硫化合物,来自于十字花科蔬菜(如西兰花芽苗)中发现的硫代葡萄糖苷前体-萝卜硫苷(一种硫代葡萄糖苷或b-硫代葡萄糖苷-N-羟基硫酸盐)(图1)(Zhang et al., 1992; Fahey et al., 1997; Kwak and Kensler, 2010; Kensler et al., 2013; Fahey et al., 2015)。众所周知,通过内源性黑芥子酶(Fahey et al., 2015),萝卜硫苷可以转化为SFN。SFN的有益作用被认为是通过激活Nrf2通路,再经ARE上调II相解毒酶和抗氧化蛋白来介导的(Suzuki et al., 2013a; Suzuki and Yamamoto, 2015)。
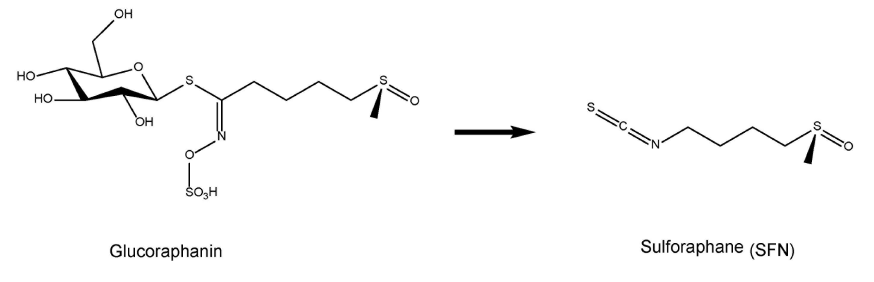
图1 萝卜硫素及其前体萝卜硫苷的化学结构。十字花科蔬菜含有萝卜硫苷,一种萝卜硫素(SFN)的硫代葡萄糖苷衍生物)。
TBE-31 [(±)-(4βS,8αR,10αS)-10α-乙炔基-4β,8,8-三甲基-3,7-二氧代-3,4β,7,8,8α,9,10,10α-八氢萘-2,6-二腈]和MCE-1 [(±)-3-乙炔基-3-甲基-6-氧代环己基-1,4-二烯腈]是新型的Nrf2激动剂(Honda et al., 2007; Dinkova-Kostova et al., 2010; Honda et al., 2011; Kostov et al., 2015; 图2)。富马酸二甲酯(图2)是一种治疗多发性硬化症的新型口服药物,通过Nrf2依赖性抗氧化反应起到神经保护作用(Al-Jaderi and Maghazachi, 2016; Mills et al., 2018)。甲基巴多酮是2-氰基-3,12-二氧代齐墩果-1,9-二烯-28-酸(CDDO)的C-28甲酯,称为CDDO-ME(图2),是合成三萜类化合物的衍生物。甲基巴多酮已用于治疗癌症(包括白血病和实体瘤)、慢性肾病和其他疾病(Wang et al., 2014)。在日本正在进行甲基巴多酮治疗糖尿病肾病的临床试验(Yamamoto et al., 2018),尽管在美国由于终末期肾病患者心脏并发症的发生而暂停了其临床试验(Pergola et al., 2011; de Zeeuw et al., 2013)。
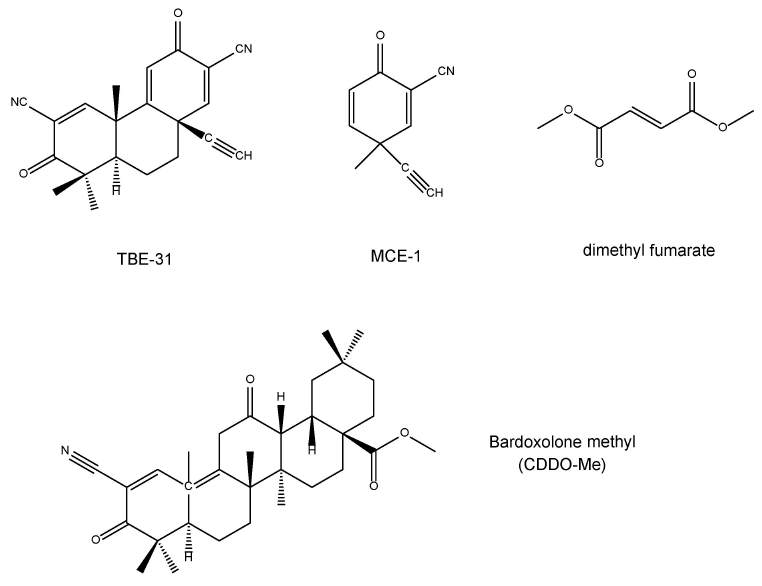
图2 Nrf2激动剂的化学结构(TBE-31,MCE-1,富马酸二甲酯,甲基巴多酮)。
Nrf2激动剂对轴突生长的影响
神经可塑性,包括轴突生长和神经保护,在细胞水平上对治疗药物的有益效果起着至关重要的作用(Lu and Dwyer, 2005;Williams and Dwyer, 2009)。Yao等(2016b)报告,在PC12细胞中,SFN增加了轴突生长的细胞的数量。此外,Nrf2 siRNA处理可阻断SFN对NGF(神经生长因子)诱导的轴突生长的增强作用,但不能阻断阴性对照(Yao et al., 2016bb),表明SFN通过激活Nrf2增强了NGF诱导的轴突生长。
Yao 等(2016a)报道,TBE-31和MCE-1也增强了NGF诱导的PC12细胞的轴突生长。Nrf2 siRNA阻断了TBE-31和MCE-1对PC12细胞轴突生长的潜在影响。另有报道,黄芪皂苷IV通过激活Akt依赖的Nrf2通路减弱了铅诱导的轴突生长抑制(Yu et al., 2017)。总之,Nrf2激动剂可通过激活Nrf2来促进轴突的生长(Yang et al., 2015c; Yao et al., 2016a,b)。
具有抑郁样表型啮齿动物Keap1-Nrf2信号转导的变化
慢性社会挫败应激(CSDS)模型被广泛用作抑郁症的动物模型(Krishnan and Nestler, 2008; Nestler and Hyman, 2010; Golden et al., 2011)。CSDS后抑郁样表型的易感小鼠血液中促炎细胞因子水平较高[例如肿瘤坏死因子(TNF)-α、白细胞介素(IL)-6、IL-10和IL-1B](Zhang et al., 2017a)。
Western blot分析显示,抑郁样表型小鼠的CA3区、DG和PFC中的Keap1和Nrf2蛋白水平显著低于对照小鼠(Yao et al., 2016b)。相反,与对照组相比,CA1区和伏隔核(NAc)中的Keap1和Nrf2蛋白水平没有差异(Yao et al., 2016b)。这些发现表明,CA3、DG和PFC中较低水平的Keap1和Nrf2可能与CSDS后的抑郁样表型有关。
习得性无助(LH)模型也被用作抑郁症的动物模型(Krishnan and Nestler, 2008)。在LH模型中,大约20-40 %的大鼠对不可避免的应激有弹性(Yang et al., 2015a,b; Yang et al., 2016)。LH(易感)大鼠的血清(原文血液,对照参考文献确定为血清)IL-6水平高于对照组和弹性大鼠(而mPFC组织中IL-6水平无显著性差异,译者注)(Yang et al., 2015a),这表明外周炎症可能有助于应激弹性,而不是对应激的易感性。LH(易感)大鼠海马PFC和DG中的Keap1和Nrf2蛋白水平低于对照组和非LH(弹性)大鼠(Zhang et al., 2018)。这些结果表明,Keap1-Nrf2信号可能有助于应激弹性,这与主要精神疾病的病理生理学有关(Zhang et al., 2018)。
严重抑郁症患者样本中Keap1-Nrf2信号转导的变化
与健康对照组(n=35)相比,严重抑郁症(MDD)患者(n=30)的Nrf2及其调节因子Keap1,以及外周血单核细胞胞浆中的NF-kB表现出较高水平,这表明抑郁症可能以转录因子Keap1-Nrf2的上调为特征(Lukic et al., 2014)。利用全基因组转录谱和基于启动子的生物信息学策略,Mellon等人(2016)测量未服用药物的MDD受试者(n=20)及其年龄和性别匹配的健康对照组(n=20)的白细胞中白细胞转录因子(TF)活性。在非药物治疗的MDD受试者的白细胞中,生物信息学分析显示cAMP应答元件结合/激活TF(CREB/ATF)的转录活性增加,与Nrf2相关的TFs活性增加。8周的抗抑郁治疗与抑郁症状显著降低和Nrf2活性降低有关,但与CREB/ATF活性无关。相比之下,其他转录调控途径,包括核因子kappa-b细胞(NF-kB)、早期生长反应蛋白1-4(EGR1-4)、糖皮质激素受体和干扰素反应性TFs,在疾病功能或治疗方面均无差异。这些结果表明,Nrf2信号可能通过激活免疫细胞转录组动力学而促进MDD,最终可能通过循环介质影响动机和情感过程(Mellon et al., 2016)。
精神疾病患者的死后组织是一种未充分利用的物质,可用于转化遗传和/或临床前研究(Hashimoto et al., 2007; McCullumsmith et al., 2013; Mechawar and Savitz, 2016; Yang et al., 2017)。一项使用尸检后大脑样本的研究表明,与对照组相比,MDD和双相情感障碍患者的顶叶皮质中的Keap1和Nrf2表达降低(Zhang et al., 2018)。最近的一项研究显示,MDD患者背外侧额叶前部皮质中Nrf2的表达降低了(-21%)(Martín-Hernández et al., 2018)。这些结果表明,Keap1-Nrf2信号的降低在MDD和双相情感障碍等心境障碍的病理生理学中起着关键作用(Zhang et al., 2018)。
人类Nrf2启动子基因的单核苷酸多态性
人类中的Nrf2活性主要通过蛋白质稳定性来调节,主要由Keap1调节,但也在转录水平上调节(Yamamoto et al., 2018)。在人类中,位于转录起始位点上游617 bp处的Nrf2启动子单核苷酸多态性(SNP:rs6721961)降低了Nrf2转录水平(Yamamoto et al., 2006; Yamamoto et al., 2018;图3)。荧光素酶分析显示-617位(C变到A)的多态性影响Nrf2的基础水平,从而导致ARE介导基因的转录减弱(Marzec et al., 2007;图3)。有趣的是,拥有这种单核苷酸多态性的受试者更易患急性肺损伤和相关疾病(Marzec et al., 2007),而且这种单核苷酸多态性也被发现与非小细胞肺癌的发病率相关(Suzuki et al., 2013b)。此外,还报告了这种单核苷酸多态性的种族差异(Marzec et al., 2007)。因此,研究这种单核苷酸多态性是否会影响对心境障碍的易感性是有意义的。
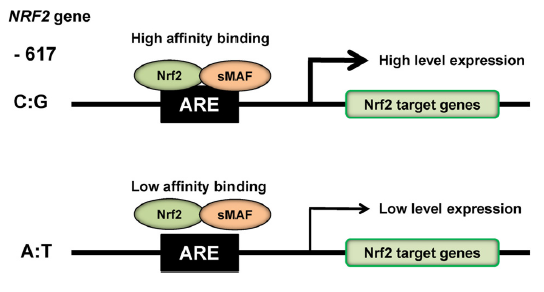
图3 Nrf2基因启动子区的单核苷酸多态性(rs6721961)。Nrf2基因启动子的单核苷酸多态性(rs6721961)改变了Nrf2基因的转录水平,导致Nrf2靶基因的表达发生改变。Nrf2/sMAF(small MAF)蛋白复合物通过占据包含抗氧化反应元件(ARE)的顺式作用增强子序列来调节氧化应激反应。Yamamoto等人的轻微修改版本(2018)。
Nrf2-KO小鼠的抑郁样表型
Nrf2-KO小鼠血清TNF-α、IL-6、IL-10和IL-1b水平显著高于野生型小鼠,提示Nrf2-KO小鼠有炎症(Yao et al., 2016b)。在悬尾实验(TST)和强迫游泳试验(FST)中,Nrf2-KO小鼠的TST和FST静止时间高于WT小鼠。在1%蔗糖偏好实验(SPT)中,Nrf2-KO小鼠的蔗糖偏好低于WT小鼠,说明Nrf2-KO小鼠快感缺乏。与此同时,Nrf2-KO小鼠的CA3区、DG和PFC中的脑源性神经营养因子(BDNF)及其受体TrkB信号低于WT小鼠。此外,KO小鼠CA3区、DG和PFC中AMPA受体1(GluA1)和突触后密度蛋白95(PSD-95)的蛋白水平低于WT小鼠。有趣的是,7,8-二羟基黄酮(一种TrkB激动剂)通过刺激PFC、CA3区和DG中的TrkB,在Nrf2-KO小鼠中产生抗抑郁作用(Yao et al., 2016b)。此外,抗炎药罗非昔布逆转了Nrf2-KO小鼠的抑郁样行为(Martín-de-Saavedra et al., 2013)。另外,选择性5-羟色胺再摄取抑制剂(SSRI)氟西汀的长期治疗增加了皮质酮(CORT)处理的Nrf2-KO小鼠皮质和海马中的BDNF(Mendez-David et al., 2015),表明恢复Nrf2信号传导仅是氟西汀诱导的神经保护作用中的一部分(文献中说明Nrf2-KO小鼠丢失Nrf2会导致慢性炎症引起抑郁样表型,Nrf2信号与抑郁样表型;氟西汀可逆转CORT诱导的小鼠焦虑/抑郁样行为(焦虑/抑郁小鼠模型,CORT模型),恢复CORT诱导的皮质和海马区Nrf2蛋白水平及其靶基因的降低,译者注)。这些研究结果表明,Nrf2通过有效的抗炎作用在啮齿动物抑郁样表型中起着关键作用。总之,Nrf2-KO小鼠很可能通过炎症、BDNF-Trkb信号下降和突触生成减少表现出抑郁样表型(图4,原文为图5,译者注;Yao et al., 2016b)。
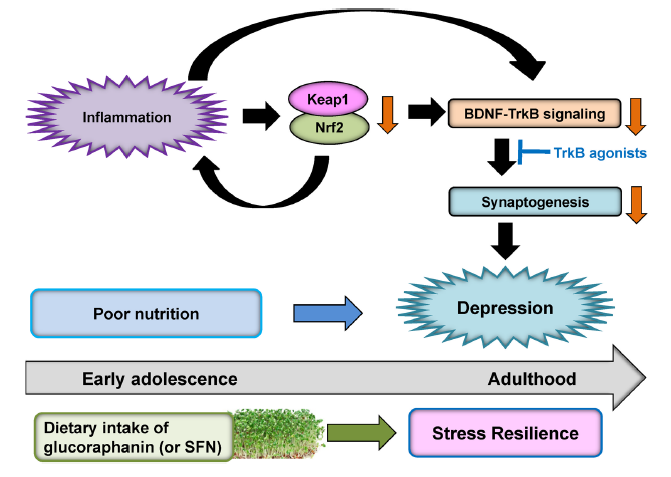
图4 提出的Keap1-Nrf2系统在抑郁症中作用的假说。炎症导致前额皮层和海马中Keap1和Nrf2表达降低。随后,炎症诱导的Keap1和Nrf2蛋白减少可导致BDNF-TrkB信号下降和突触发生减少,导致抑郁样表型。TrkB激动剂可能具有抗抑郁作用。青春期早期摄入十字花科蔬菜中的萝卜硫苷(或SFN)可能会在成年时授予应激弹性,而营养不良可能在应激或炎症引起的抑郁发病中起作用。
相反,Bouvier等(2017)报告称,Nrf2-KO小鼠没有表现出抑郁样表型,尽管KO小鼠以氧化应激和海马CA3区锥体细胞的解剖改变为特征。然而,当暴露于慢性轻度应激下3周时,Nrf2-KO小鼠出现了抑郁样表型,该过程可通过用抗氧化剂预处理预防(Bouvier et al., 2017)。这项研究也表明了Nrf2依赖的持续氧化应激在应激诱导的抑郁症易感性中的作用。
Nrf2激动剂在啮齿动物抑郁模型中的抗抑郁作用
当对啮齿类动物施用细菌内毒素脂多糖(LPS)时,炎症发生后24小时观察到抑郁样行为(Dantzer et al., 2008; O’Connor et al., 2009; Zhang et al., 2014; Zhang et al., 2016)。以如SSRIs和5-羟色胺和去甲肾上腺素再摄取抑制剂(SNRIs)的抗抑郁药预处理后,可预防由LPS诱导的抑郁样行为和血清促炎细胞因子如TNF-α的改变(Ohgi et al., 2013; Yao et al., 2015; Dong et al., 2016)。所有这些发现都表明炎症与抑郁症相关,抗炎药可以改善抑郁症患者的抑郁症状。
SFN预处理显著阻断了单次LPS给药后血清TNF-α水平的升高(Zhang et al., 2017b)。此外,SFN显著增强了LPS给药后血清IL-10的升高。LPS给药后,SFN减弱了TST和FST静止时间的延长。另外,SFN恢复LPS诱导的脑区蛋白(如BDNF、PSD-95和GluA1)和树突密度到对照水平(Zhang et al., 2017b)。此外,TBE-31或MCE - 1可降低LPS给药后TNF-α水平的升高。给予TBE-31或MCE-1可减少LPS给药后TST和FST静止时间的增加(Yao et al., 2016a)。
在CSDS模型中,SFN预处理减弱了社交回避时间的降低和蔗糖偏好的下降。此外,SFN还可减弱了抑郁样表型小鼠PFC和海马中Nrf2和Keap1蛋白水平的下降(Yao et al., 2016b)。Li等(2018)报告称,PFC、海马和骨骼肌中Keap1-Nrf2信号的降低可能促成了保留性神经损伤(SNI)后快感缺乏的易感性,SFN通过恢复Keap1-Nrf2信号的正常对SNI大鼠发挥有益作用。这些结果表明,Keap1-Nrf2信号在抑郁症中起重要作用,SFN是一种能够刺激Keap1-Nrf2信号通路的预防性化合物(Yao et al., 2016b; Zhang et al., 2017b; Li et al., 2018)。综上所述,如SFN、TBE-31和MCE-1等Nrf2激动剂可能是治疗炎症(或应激)相关抑郁症的潜在预防或治疗性药物(Yao et al., 2016a,b; Zhang et al., 2016, 2017b; Li et al., 2018)。
CSDS抑郁模型中膳食摄入SFN前体的影响
SFN是由它的前体-十字花科蔬菜中萝卜硫苷在体内产生的。在此之前,我们证明,在少年期和青年期的饮食中摄入富含0.1%萝卜硫苷的食物可以预防成年时苯环己哌啶(原为一种兽用镇静剂,又称“天使粉”,是一种非法的迷幻药物,译者注)诱导的认知缺陷和PFC中的小清蛋白(PV)阳性细胞的丢失(Shirai et al., 2015)。此外,我们还报告,在少年期和青年期的饮食中摄入富含0.1%萝卜硫苷的食物可预防母体免疫激活后的成年后代发生精神病(Matsuura et al., 2018)。这些发现表明,在高危精神病受试者中,饮食中摄入富含萝卜硫苷的蔬菜可能会预防年轻成人向精神病患者的转变(Hashimoto, 2014; Shirai et al., 2015; Matsuura et al., 2018)。
有趣的是,在少年和青年阶段,饮食中摄入含0.1%萝卜硫苷的食物可以预防成人期CSDS后的抑郁样表型(Yao et al., 2016b)。因此,在少年期和青年期的饮食中摄入含0.1%萝卜硫苷的食物可以赋予其成年期应激弹性。
健康受试者和神经发育障碍患者中SFN的临床研究
Sedlak等(2018)报告称,在每天SFN口服给药7天后,健康受试者的血液谷胱甘肽(GSH)水平上升。此外,观察到血液与丘脑中的GSH水平在SFN治疗后和治疗前的比率存在显著正相关(个体GSH(处理后血液)/GSH(处理前血液)与GSH(处理后脑部)/GSH(处理前脑部)显著正相关,译者注),且治疗后大脑GSH水平持续升高。这项研究表明了探索外周GSH与临床/神经心理测量之间关系的价值,以及SFN对神经精神疾病中功能改变测量的影响。
一项随机、双盲、安慰剂对照研究表明,富含SFN的西兰花芽苗提取物可以改善患有自闭症谱系障碍的青年男性受试者的社交互动、异常行为和言语交流(Singh et al., 2014; Lynch et al., 2017)。此外,一项初步研究表明,,补充富含萝卜硫苷的西兰花芽苗提取物8周对治疗精神分裂症患者的认知障碍是有效的(Shiina et al., 2015))。总之,SFN可能是治疗神经发育障碍的潜在化合物。
Nrf2在其他潜在化合物的抗抑郁机制中的作用
西洛他唑用于治疗周围血管疾病患者间歇性跛行症状。在慢性约束应激(CRS)模型中,西洛他唑可预防抑郁样行为(Abuelezz and Hendawy, 2018)。此外,西洛他唑还调节了CRS大鼠海马中Nrf2蛋白、血红素加氧酶-1(HO-1)和NAD(P)H:醌氧化还原酶-1(NQO-1)基因的表达。这些发现表明,西洛他唑通过刺激Nrf2通路介导的氧化还原防御机制来预防氧化应激,从而具有预防性抗抑郁作用(Abuelezz and Hendawy, 2018)。
从芹菜籽中提取的小分子化合物DL-3-N-正丁基苯酞(NBP)经中国国家食品药品监督管理局批准用于治疗缺血性中风(Abdoulaye and Yi, 2016)。NBP减弱了大鼠的抑郁样行为,增加了促炎细胞因子(如IL-1b和IL-6)的表达。除抗炎作用以外,NBP还降低了LPS诱导的海马氧化应激反应,增强了Nrf2靶向信号(Yang et al., 2018)。
一项随机、双盲、安慰剂对照试验表明,NBP与安慰剂相比对阿尔茨海默病评估量表认知分量表(ADAS-cog)和临床医生基于访谈的总体印象变化加护理者输入(CIBIC-plus)的影响更大。NBP相关不良事件不常见,主要由轻度胃肠道症状组成(Jia et al., 2016)。因此,研究NBP是否能改善抑郁症患者的抑郁症状具有重要意义。
Nrf2抑制剂
与Nrf2激动剂相比,Nrf2抑制剂的开发尚处于起步阶段(Yamamoto et al., 2018)。例如,Nrf2持续激活的癌症表现出对Nrf2的耐药性和细胞增殖功能的高度依赖性(Yamamoto et al., 2018)。植物性产品鸦胆子苦醇(图5,原文为图4,下同,译者注)降低了Nrf2蛋白水平,并使癌细胞对化疗和放疗敏感(Ren et al., 2011)。另一种Nrf2抑制剂常山酮(图5)是常山乙素的一种卤代合成衍生物,常山乙素是一种天然的喹唑啉酮生物碱,在中草药常山(Dichraa febrifuga)中发现。常山酮对具有构成性Nrf2稳定性的癌细胞起化疗增敏作用(Tsuchida et al., 2017)。此外,Singh等(2016)证明ML-385 [N-[4-[2,3-二羟基-1-(2-甲基苯甲酰)-1H-吲哚-5-基]-5-甲基-2-噻唑基]-1,3-苯并二茂-5-乙酰胺](图5)是一种新颖的、特异性的Nrf2抑制剂。因此,研究这些Nrf2抑制剂是否能影响啮齿类动物的抑郁样表型具有重要意义。
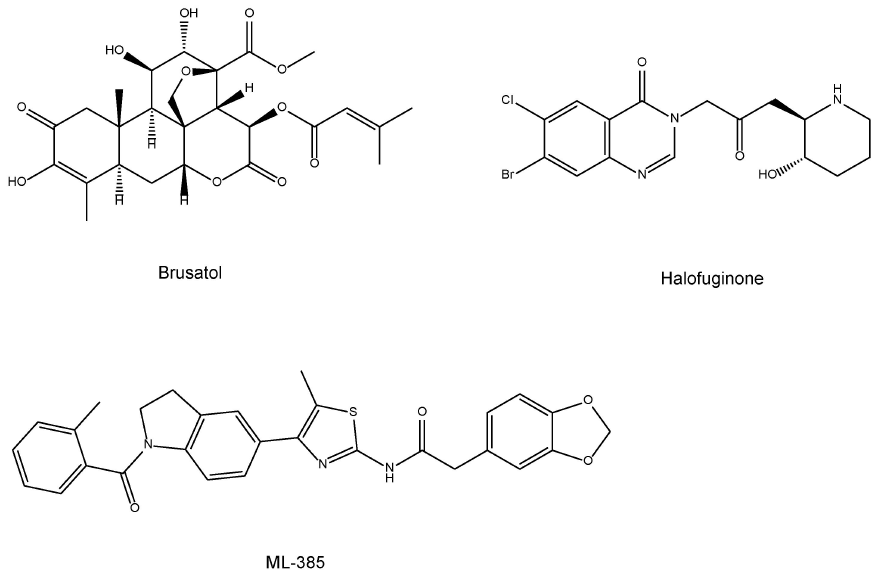
图5 Nrf2抑制剂的化学结构(鸦胆子苦醇、常山酮、ML-385)。
结论与展望
具有抑郁样表型的啮齿动物血液中促炎细胞因子水平较高,表明炎症在啮齿动物抑郁样表型中起作用。此外,具有抑郁样表型的啮齿动物在PFC和海马体中的Keap1和Nrf2表达较低(Yao et al., 2016b; Zhang et al., 2017b)。有趣的是,我们发现MDD和双相情感障碍患者的顶叶皮层中Keap1和Nrf2的表达降低(Zhang et al., 2018)。考虑到BDNF-Trkb信号在抑郁症中的重要作用(Nestler et al., 2002; Hashimoto et al., 2004; Duman and Monteggia, 2006; Hashimoto, 2010; Zhang et al., 2016),炎症(或应激)诱导的Keap1-Nrf2系统的减少可能会减少BDNF-Trkb信号和突触生成,导致抑郁样表型(图4,原文为图5,译者注)。值得注意的是,Trkb激动剂7,8-DHF对Nrf2-KO小鼠(Yao et al., 2016b)、LPS处理小鼠(Zhang et al., 2014)和CSDS敏感小鼠(Zhang et al., 2015)具有抗抑郁作用,这表明Keap1-Nrf2系统与BDNF-Trkb信号传导之间可能存在联系(Mendez-David et al., 2015; Yao et al., 2016b; Zhang et al., 2017b; Li et al., 2018; 图4,原文为图5,译者注)。
青年期早期的营养状况可能对成年期精神疾病的发病和严重程度有很大影响(Paus et al., 2008; O’Connor and Cryan, 2014)。在过去的十年中,人们对早期干预精神疾病(如抑郁症)的潜在益处越来越感兴趣(Paus et al., 2008; O’Connor and Cryan, 2014; Sarris et al., 2015; Correll et al., 2018)。
临床前研究结果表明,在少年和青年期的饮食中摄入萝卜硫苷可以预防CSDS或LPS给药后出现的抑郁样行为(Yao et al., 2016b; Zhang et al., 2017b),表明萝卜硫苷对抑郁症有预防作用。因此,在少年和青年期饮食中摄入萝卜硫苷(或SFN)可能会在成年时带来应激弹性(图4)。因此,在儿童和青少年阶段的饮食中摄入萝卜硫苷(或SFN),有可能预防在成年期出现抑郁症。由于心境障碍患者的复发率较高,因此饮食中摄入萝卜硫苷(或SFN)可预防或减少抑郁症患者因炎症和/或应激引起的复发。
本文由福山生物整理翻译,转载请注明出处。