






最新资讯



-
电话: 400-113-6988 邮箱: dongfangxicao@163.com 地址: 深圳市南山区粤海街道高新中三道9号环球数码大厦19楼
↓ 微信扫码识别关注我们 ↓ 
运用系统医学方法阐明转录因子NRF2作为慢性疾病的治疗靶标
发表于:2019-05-14 作者:admin 来源:本站 点击量:28684
Antonio Cuadrado et al., PHARMACOLOGICAL REVIEWS, 2018
目 录
缩写:
AD: 阿尔茨海默病; ALS: 肌萎缩侧索硬化症; AMPK,AMP活化蛋白激酶; ARE: 抗氧化反应元件; β-TrCP: 含有E3泛素蛋白连接酶的β-转导蛋白重复序列; bZip: 碱性区域 - 亮氨酸拉链; CDDO: 2-氰基-3,12-二氧代-油酸-1,9(11)- 二烯-28-酸; CDDO-Me: CDDO-甲酯; COPD:慢性阻塞性肺病; CUL3,Cullin 3; DC:树突状细胞; DMF: 富马酸二甲酯; EAE:实验性自身免疫性脑脊髓炎; ECH:具有Cap'n'collar同源性的红系细胞衍生蛋白; GCLC:g-谷氨酰半胱氨酸连接酶催化亚基; GCLM:g-谷氨酰半胱氨酸连接酶调节剂亚基; GI:胃肠道; GSH:谷胱甘肽; GSK-3:糖原合成酶激酶3; HFD,高脂饮食; HNSCC:头颈部鳞状细胞癌; HO-1,血红素加氧酶-1; IBD:炎症性肠病; IκB:κ-B抑制剂; IKK:IκB激酶; IL:白细胞介素; KEAP1:kelch样ECH相关蛋白1; LDL:低密度脂蛋白; LPS:脂多糖; MAF:肌肉腱膜纤维肉瘤蛋白; MMF:富马酸单甲酯; MS:多发性硬化; NASH:非酒精性脂肪性肝炎; NF-κB:活化B细胞核因子κ-轻链增强子的p65亚基; NQO1:NADPH:醌氧化还原酶; NRF2:核因子(红细胞衍生的2)样2; PD:帕金森病; PI3K:磷脂酰肌醇3-激酶; PPI:蛋白质-蛋白质相互作用; PTEN:磷酸酶和张力蛋白同系物; RA: 类风湿性关节炎; RBX1:RING-box蛋白1; RNS:活性氮物种; ROS:活性氧; SFN:萝卜硫素; SLE:系统性红斑狼疮; SNP:单核苷酸多态性; SQSTM1:sequestosome 1; STAT:信号转导和转录激活因子; T2DM:2型糖尿病; TGF:转化生长因子; Th,T辅助子; TNF:肿瘤坏死因子; Treg:T调控。
Antonio Cuadrado et al., PHARMACOLOGICAL REVIEWS, 2018
摘要
系统医学是基于疾病机理,而非基于具体症状或器官的疾病研究方法,并以非假设驱动的方式识别治疗靶标。本论文中,我们将系统医学方法应用于转录因子核因子( NRF2)的研究,通过交叉验证其在蛋白质-蛋白质相互作用网络(NRF2相互作用组)中的作用,该相互作用网络在功能上与低水平应激、慢性炎症、代谢改变和活性氧形成有关的细胞保护相关联。这些分子谱的多尺度网络分析表明,NRF2表达和活性的改变是疾病子网路中的常见机制(NRF2病变)。该相互作用网络关联了明显的异质性表型,如自身免疫、呼吸、消化、心血管、代谢和神经退行性疾病,以及癌症。重要的是,系统医学方法在计算机上匹配并证实了在临床开发的不同阶段在体内验证的NRF2调节药物的若干应用。从药理学上看,这些药物的特征多样,有亲电的富马酸二甲酯、合成的三萜类化合物如巴多索酮甲基和萝卜硫素、蛋白-蛋白或DNA -蛋白相互作用抑制剂,甚至有注册的药物如二甲双胍和他汀类药物,它们可以激活NRF2并且可能被重新用于指示 NRF2疾病表型群。因此,NRF2代表了经典和系统医学方法充分接受的第一类目标之一,这些方法通过关注一组似乎具有机械联系的疾病表型来促进药物开发和药物新用。 因此,由此产生的NRF2药物组可能会迅速为这类慢性疾病提供一些令人惊讶的临床选择。
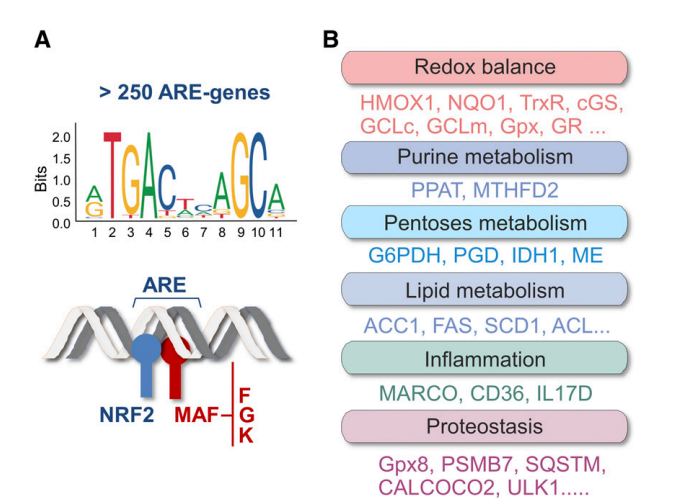
图1. NRF2作为细胞保护应答的主要调节因子。 (A)NRF2通过其bZip结构域与MAF家族成员异二聚体化。异二聚体与称为ARE的增强子序列结合,所述增强子序列存在于超过250个基因(ARE基因)的调节区中。 (B)如所示,这些基因参与氧化还原代谢,炎症和蛋白质稳态平衡的控制。 NFE2L2中易感性SNPs的存在,脑部尸检中其靶基因水平的升高以及临床前研究的阳性数据表明,NRF2激活可能抵消蛋白质稳态,氧化还原和炎症控制的不平衡。 AC1: ATP柠檬酸裂解酶; ACC1: 乙酰辅酶A羧化酶1; CALCOCO2: 钙结合和卷曲螺旋结构域2; cGS: c-谷氨酸半胱氨酸合成酶; FAS: 脂肪酸合成酶; G6PDH: 葡萄糖-6-磷酸脱氢酶; Gpx: 谷胱甘肽过氧化物酶; Gpx8: 谷胱甘肽过氧化物酶8; GR: 谷胱甘肽还原酶; HMOX1: 血红素加氧酶-1; IDH1: 异柠檬酸脱氢酶1; ME: 苹果酸酶; MTHFD2: 亚甲基四氢叶酸脱氢酶2; PGD: 磷酸葡萄糖酸脱氢酶; PPAT: 磷酸核糖焦磷酸酰胺转移酶; PSMB7: 蛋白酶体亚基b-7型; SCD1: 硬脂酰辅酶A去饱和酶; TrxR: 硫氧还蛋白还原酶; ULK1: unc-51像自噬激活激酶1。
NRF2在临床上的重要意义在于它可能具有药理学靶向性,对患者有益。调节NRF2转录活性的主要机制是通过E3连接酶适配器KEAP1(具有Cap'n'collar同源性(ECH)的Kelch样红系细胞衍生蛋白 - 相关蛋白1)来控制蛋白的稳定(图2)。KEAP1是同型二聚体蛋白,其将NRF2与Cullin 3和RING-box蛋白1(CUL3 / RBX1)形成的E3连接酶复合物连接。在稳态条件下,KEAP1同二聚体的N-末端结构域在两个氨基酸序列上与一个分子的NRF2结合,具有低(天冬氨酸,亮氨酸和甘氨酸; DLG)和高(谷氨酸,苏氨酸,甘氨酸和谷氨酸; ETGE)亲和力,因此,通过CUL3 / RBX1(Tong等人,2007)将NRF2呈现为泛素化,并随后通过蛋白酶体降解。KEAP1是一种氧化还原和亲电子传感器,在关键半胱氨酸修饰后失去其抑制NRF2的能力(图2;生物标志物作为NRF2特征并参与监测目标)。调节NRF2稳定性的另一种机制是由糖原合成酶激酶3(GSK-3)介导的磷酸化(图2)。这种激酶磷酸化NRF2(天冬氨酸,丝氨酸,甘氨酸,异亮氨酸,丝氨酸; DSGIS)的结构域,因此为含有E3泛素蛋白连接酶(b-TrCP)的E3连接酶接头b-转导蛋白重复序列产生识别基序,该基序将NRF2呈递给 CUL1 / RBX1复合物,导致NRF2泛素依赖性蛋白酶体降解的替代途径。因此,KEAP1和GSK-3 / b-TrCP分别在氧化还原稳态和细胞信号传导的情况下严格控制NRF2蛋白水平(Cuadrado,2015)。已经报道了NRF2在蛋白质,mRNA或基因水平上调节的其他机制(Hayes and Dinkova-Kostova,2014),但至少这两种机制适合于药理学调节。
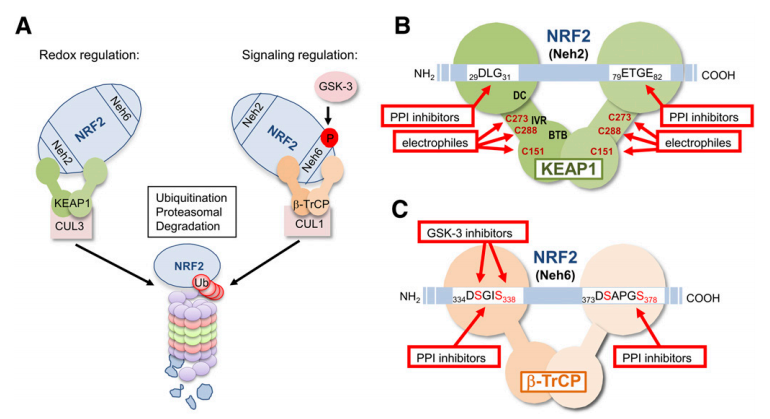
图2. KEAP1和β-TrCP对NRF2稳定性的调节及其药理学靶向。(A)根据双重调节模型(Rada等,2011),NRF2的两个结构域,称为Neh2和Neh6,参与NRF2降解分别响应氧化还原和亲电子变化(KEAP1)和信号激酶(β-TrCP)。Neh2结构域结合E3连接酶适配子KEAP1,其呈递NRF2用于泛素化至CUL3 / RBX1复合物。 Neh6结构域需要GSK-3的先前磷酸化以结合E3连接酶适配子β-TrCP并随后通过CUL1/ RBX1复合物进行泛素化(详见文本)。(B)NRF2和KEAP1之间结合的细节以及当前针对这种相互作用的策略。KEAP1同型二聚体在Neh2结构域的两个基序处结合NRF2:低亲和力(29-DLG-31)和高亲和力(79-ETGE-82)结合位点。目前破坏这种相互作用的策略包括:改变半胱氨酸C151,273和288的巯基的亲电试剂; 改变NRF2与KEAP1的DC结构域的对接的PPI抑制剂。(C)NRF2和β-TrCP的假设结合,并提出了针对这种相互作用的策略。当它被GSK-3(Rada等,2011,2012)磷酸化时,β-TrCP同型二聚体在磷酸基序334-DSGIS-338处与Nhe6结构域结合,当与磷酸基序373-DSAPGS-378结合时独立于GSK-3(Chowdhry等,2013)。在该图中,我们假设,与KEAP1类似,一个β-TrCP同二聚体在两个磷酸基序上与一个NRF2分子相互作用,但仍缺乏实验证据。破坏这种相互作用的两种可能策略包括使用GSK-3抑制剂和PPI抑制剂。
网络医学的最新进展提供了定量工具来表征基因与其相互作用(相互作用组)之间的相互作用如何与病理学相关(Barabasi等,2011; Vidal等,2011; Guney等,2016),分子网络失调在各种疾病中如何普遍(Menche等,2015),以及疾病如何在特定组织中表现(Kitsak等,2016)。为了从系统医学的角度理解NRF2在病理学中的相关性,首先我们已经生成了人类相互作用组图。我们整合并策划了与NRF2调节途径相关的蛋白质之间的物理相互作用的信息(Hayes and Dinkova-Kostova,2014; Cuadrado,2015)。交互数据来自最近发表的人类相互作用组,其汇编了几种蛋白质 - 蛋白质相互作用(PPI)资源的数据(Turei等,2013; Menche等,2015)。然而,目前可用于NRF2相互作用组开发的信息受限于以下事实:因为NRF2是非常短的半衰期蛋白质,所以可能未检测到一些有意义的相互作用。然而,在相互作用组中发现了一些与NRF2物理性相互作用的众所周知的蛋白质,包括KEAP1,b-TrCP和MAF。另一组NRF2相互作用蛋白对应于具有调节基因表达功能的核蛋白。这些包括与bZip转录因子,核受体或共激活因子相关的蛋白质,或涉及组蛋白乙酰化的蛋白质。因此,NRF2相互作用组证明除了与该转录因子直接相关的基因调控之外的其他基因调控机制。NRF2在几个残基处被磷酸化,因此预期它与几种激酶相互作用,所述激酶包括GSK-3和几种蛋白激酶C同种型。这些激酶是一些膜受体和衔接子/支架蛋白的下游。除了这种物理相互作用,我们已经确定了NRF2邻域中富集的几种生物功能,包括代谢过程,如戊糖,四吡咯,血红素,葡萄糖6-磷酸,半胱氨酸,GSH,甘油醛-3-磷酸和NADPH的生物合成。大多数这些调节蛋白不直接相互作用,而是通过作为介质的蛋白质连接(图3;特定相互作用分子的更详细描述可在这个网址找到http://sbi.imim.es/data/nrf2/)。
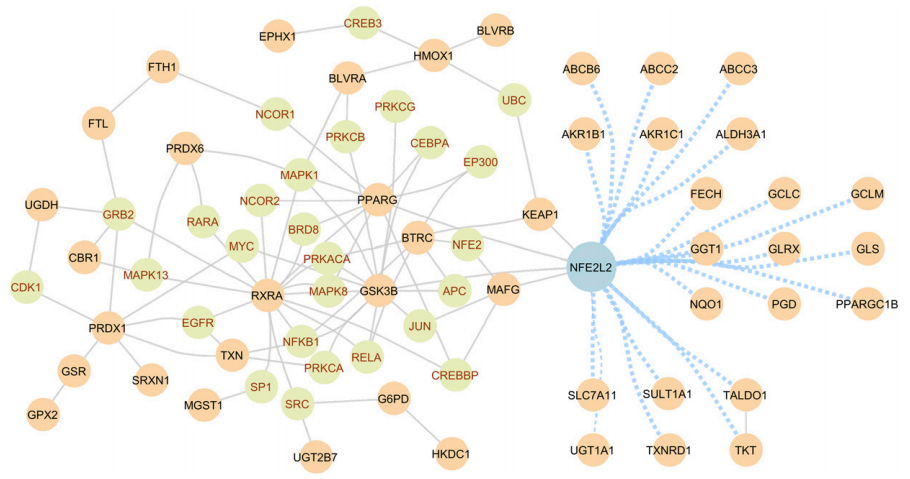 图3. 在人类相互作用组中定位NRF2调节途径。NRF2通过协调各种蛋白质的活性在病理性ROS形成以及炎症和代谢反应中起关键作用。这些相互作用构成分子相互作用网络,即NRF2相互作用组。参与NRF2调节途径的蛋白质之间的已知物理相互作用(即调节蛋白:棕色圆圈)和它们连接的蛋白质(即,介体蛋白质:绿色圆圈)用灰色连接显示。涉及多于一种介体蛋白连接到NRF2的调节蛋白用蓝色连接显示。调节蛋白质来自文献,它们的相互作用来自各种资源,包括IntAct,MINT,BioGRID,HPRD,KEGG和PhosphoSite。可在线获取人类相互作用组中与NRF2相互作用的所有蛋白质列表,网址为http://sbi.imim.es/data/nrf2/。
图3. 在人类相互作用组中定位NRF2调节途径。NRF2通过协调各种蛋白质的活性在病理性ROS形成以及炎症和代谢反应中起关键作用。这些相互作用构成分子相互作用网络,即NRF2相互作用组。参与NRF2调节途径的蛋白质之间的已知物理相互作用(即调节蛋白:棕色圆圈)和它们连接的蛋白质(即,介体蛋白质:绿色圆圈)用灰色连接显示。涉及多于一种介体蛋白连接到NRF2的调节蛋白用蓝色连接显示。调节蛋白质来自文献,它们的相互作用来自各种资源,包括IntAct,MINT,BioGRID,HPRD,KEGG和PhosphoSite。可在线获取人类相互作用组中与NRF2相互作用的所有蛋白质列表,网址为http://sbi.imim.es/data/nrf2/。
已经报道了几种疾病中NRF2相互作用组的扰动。我们基于DisGeNET (Pinero et al., 2017)和GeneCards (Stelzer et al., 2016)数据库,结合部分动物模型研究的知识,整理了37种nrf2相关疾病的列表,绘制了疾病组图。我们还使用DisGeNET,OMIM和GWAS数据库检索了这些病症的疾病 - 基因关联(Menche等,2015)(表1)。NRF2与每种NRF2相关疾病表型的已知疾病基因的基于相互作用组的接近度(Guney和Oliva,2014)显示在图4中。与随机选择的蛋白质相比,NRF2与消化系统和癌症(如前列腺,肝脏和肺肿瘤)的已知疾病基因显着接近,突出了NRF2在这些病理中的关键功能作用。此外,发现NRF2与代谢和心血管疾病相关的各种蛋白质均有近端表达,例如糖尿病,高血糖症,局部缺血,大脑中动脉梗塞和动脉粥样硬化。NRF2的蛋白质相互作用还将其与呼吸系统疾病相关的基因(如哮喘,肺纤维化和肺气肿)以及神经退行性疾病(如阿尔茨海默病(AD),帕金森病(PD)和肌萎缩侧索硬化症(ALS))联系起来。
表1.有NRF2相关证据的疾病群集
从DisGeNet数据库中选择具有与NRF2遗传相关的证据的疾病表型。DisGeNet整合来自各种资源的疾病基因关联信息,如UniProt,ClinVar,GWAS目录和比较毒物基因组学数据库,并根据资源数量和支持这些关联的出版物评估疾病基因关联。该清单的制定基于以下标准:1)仅选择在Pubmed中具有多于一个引用的病理表型; 2)可靠性评分设定为0.001的阈值; 3)具有非常相似的名称或重叠术语的疾病条目被简化为单个条目。
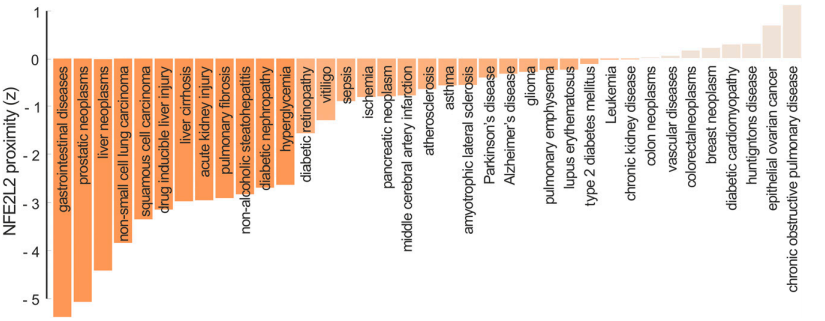
图4. 从人类相互作用组的角度看NRF2的系统医学观点。疾病是由扰乱蛋白质及其相互作用的突变引发的,影响NRF2相互作用组中的某个邻域。基于相互作用组的接近度(proximity)测量基因与那些疾病邻域的距离。 条形图显示NRF2基因(NFE2L2)与参与NRF2相关病理表型的已知疾病基因的接近程度。这些条形图强调了NRF2在消化系统疾病和癌症中的作用。对于给定的疾病,接近度首先计算从NRF2到最接近的已知疾病基因的距离,然后将该距离与使用相互作用组中随机选择的蛋白质之间的平均距离估计的随机期望进行比较。基于相互作用组的接近度报告的z分数对应于NRF2与疾病基因之间观察到的距离的显着性。负值表示观察到的距离低于偶然预期的距离。根据z得分,条纹以不同的橙色调着色:分别为近端(深橙色),近端(橙色),非近端(浅橙色)。已知的疾病基因来自DisGeNet,OMIM和GWAS目录。
因此,基于相互作用组的接近度提供了关于NRF2如何与各种病理状况相关联的观点。之前已将疾病之间的关系概括为一种网络,成为疾病组,其基于遗传(Goh等人,2007)和临床(Hidalgo等人,2009; Zhou等人,2014)的共同性来连接它们。在图5中,我们基于共享基因,症状相似性和合并症定义了疾病网络,即NRF2疾病组。NRF2似乎连接基本上由炎症过程控制的疾病,例如急性肾损伤,肝硬化和动脉粥样硬化。此外,AD,PD,亨廷顿舞蹈病和ALS等神经退行性疾病构成一个集群,与最近的研究一致,也暗示NRF2在神经炎症过程中的作用(Rojo等,2010; Lastres-Becker等,2012,2014; Jung等,2017; Wang等,2017b)。通过激酶信号转导级联对NRF2的调控(Jung et al., 2017)可以解释密切相关的癌症簇,特别是支持NRF2参与结肠和乳腺肿瘤病理性ROS形成(Gonzalez-Donquiles et al., 2017;Lu et al., 2017)。
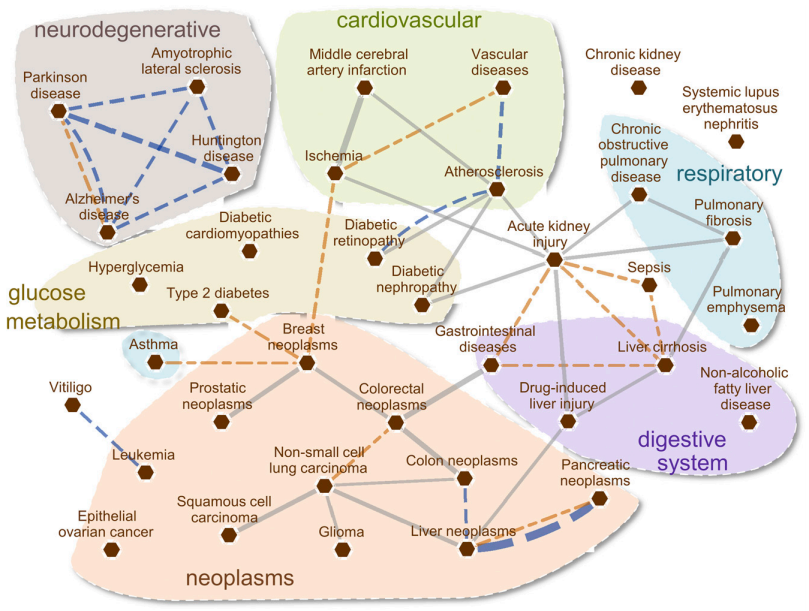

图5. NRF2疾病组的当前状态。疾病之间的关系表示为一个网络,其中病理表型由共同的遗传和临床描述符连接。图中,节点(红色六角形)代表疾病,边缘根据共同的基因、共同的症状和共病(分别为灰色、橙色和蓝色线条)表示疾病之间的相似性。与疾病相关的基因和症状被用来识别具有显著遗传和症状重叠的疾病对,这些重叠是使用Jaccard指数计算出来的。在显着的疾病 - 疾病关系中(P <0.05,通过基于观察到的基因或症状重叠的Fisher精确检验评估),仅显示具有升高的重叠和共病的链接以消除潜在的虚假连接。因此,图中包含了至少10%的疾病相关基因和超过一半的相关症状。共病信息是从医疗保险索赔中提取的,代表人群中容易同时发生的疾病对(相对风险>2)。
由于NRF2是氧化还原稳态的主要调节因子,因此它对NF-κB活性进行间接控制。脂多糖(LPS)同时激活快速、促炎性的NF-κB反应和缓慢的NRF2反应。当NRF2最大活性时,NF-κB响应随后被抑制(Cuadrado等,2014)。例如,Ras相关的C3肉毒杆菌毒素底物1(Rho家族的一个小G蛋白)激活NF-kB途径,并且NRF2过表达被阻断,而NRF2敲低增强NF-κB依赖性转录(Cuadrado等,2014)。一致地,在用LPS或肿瘤坏死因子(TNF)-α攻击的NRF2缺陷型(Nrf2-/-)小鼠中,IKK的活性恶化并导致IκB的磷酸化和降解增加(Thimmulappa等,2006a)。
NRF2还诱导抗炎表型,其调节CD8+T细胞(Sha等人,2015)以及巨噬细胞和小胶质细胞的功能(Rojo等人,2010,2014a; Brune等人,2013)。这是因为NRF2通过调节胱氨酸/谷氨酸转运蛋白和GSH合成酶?-谷氨酰半胱氨酸连接酶调节器和催化亚基[?-谷氨酰半胱氨酸连接酶调节剂亚基(GCLM)和?-谷氨酰半胱氨酸连接酶催化亚基(GCLC)]来增加巨噬细胞中的半胱氨酸和GSH水平。相反,GSH耗竭使巨噬细胞对LPS激活NRF2敏感(Diotallevi等,2017)。所有这些研究都指出NRF2是一种抗炎因子,对控制炎症反应的强度和持续时间至关重要(图6)。
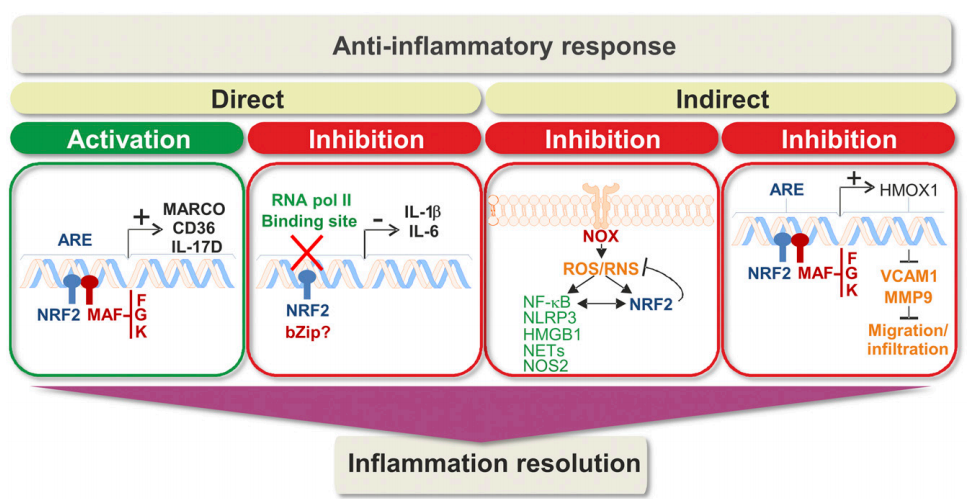
图6. 通过NRF2直接和间接调节炎症。直接作用机制包括抗炎基因的转录诱导以及促炎基因的转录抑制。 在第二种情况下,引号表示在此功能中需要进一步的工作来识别NRF2的bZip伙伴(如果有的话)。抵抗炎症的间接机制涉及ROS / RNS调节和抑制免疫细胞的迁移/浸润。总体而言,这些途径导致抗炎反应,有助于正确解决炎症。NFE2L2中多态性的存在与转录活性降低,患者中靶基因水平的改变以及来自临床前研究的有希望的数据相关,支持NRF2在炎症消退中的相关作用。
通过前馈和反馈机制产生NRF2和NF-κB串扰(图7)。在转录水平,由于NFE2L2基因的启动子区域中存在几个功能性结合位点,NF-κB激活NRF2表达,从而诱导负反馈环(Rushworth等,2012)。此外,NF-κB和NRF2转录因子都需要共激活因子CBP / p300,它是组蛋白乙酰转移酶,乙酰化并增加DNA结合能力。因此,NF-κB过表达阻碍了NRF2的CBP / p300的可用性,因此降低了其转录能力,而NF-kB敲低显示出相反的效果(Liu等人,2008)。另外,NF-κB可以促进组蛋白脱乙酰酶-3与MAF蛋白的相互作用,因此阻止它们与NRF2的二聚化(Liu等人,2008)。NF-κB将KEAP1结合并转移至细胞核,从而有利于NRF2在该细胞区室中的泛素化和降解(Yu等,2011)。 E3连接酶适配子β-TrCP标记IκBα(Winston等人,1999)和NRF2(Rada等人,2011,2012; Cuadrado,2015)用于蛋白酶体降解,因此它可以导致增加的NF-kB活性。
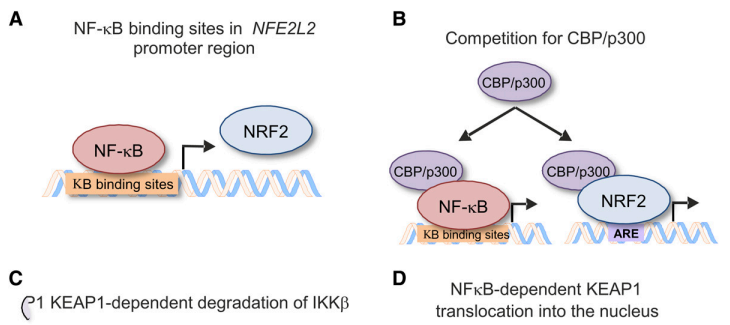


图7. NF-κB和NRF2之间的串扰发生在不同的水平。(A)已在NFE2L2的启动子区域中鉴定出响应元件。(B)NRF2和NF-κB转录因子都竞争结合转录共激活因子CREB结合蛋白(CBP / p300)。(C)NF-κB活化激酶IKKb含有ETGE基序,其允许KEAP1结合和随后的泛素蛋白-蛋白酶体降解。(D)据报道NF-κB结合并将KEAP1转移至细胞核,从而促进NRF2降解。(E)炎症过程中产生的ROS激活NF-κB和NRF2; 最后,NRF2减弱ROS并因此减弱NF-κB活性。(F)不同的促炎信号激活Rho GTP酶RAC1,导致NF-κB和NRF2活化。 然后NRF2抑制RAC1介导的NF-kB活化。
NRF2的抗炎活性被认为仅依赖于氧化还原代谢的调节或与NF-κB的串扰。然而,NRF2还可以在暴露于LPS后直接阻断巨噬细胞中促炎基因白细胞介素(IL)-6和IL-1β的转录(Kobayashi等,2016)。LPS暴露或NRF2的药理学活化导致其与这些促炎基因的近端启动子结合并阻断RNA pol II的募集。该机制似乎与NRF2与其成熟的ARE增强子的结合无关。在其他研究中,NRF2可以直接调节其他几种巨噬细胞特异性基因的表达,例如具有胶原结构的巨噬细胞受体,细菌吞噬作用所需的受体,或CD36,氧化低密度脂蛋白的清道夫受体(Harvey等,2011;Ishii和Mann,2014)。类似地,编码促炎细胞因子IL-17D的基因含有AREs,并且该NRF2-T辅助因子(Th)17轴似乎赋予针对肿瘤发生和病毒感染的保护作用(Saddawi-Konefka等,2016)。
慢性炎症过程涉及白细胞粘附到血管内皮并渗入受损组织。两种过程似乎都被NRF2与其编码血红素加氧酶-1(HO-1)的至少一种靶基因HMOX1一起调节。NRF2 / HO-1轴通过调节几种细胞粘附分子如血管细胞粘附分子1的表达来抑制炎性细胞与内皮的粘附(Banning和Brigelius-Flohe,2005; Wenzel等,2015)。另外,NRF2 / HO-1抑制巨噬细胞中的金属蛋白酶-9,这是组织内免疫细胞迁移所必需的(Bourdonnay等,2009)。
许多临床前研究报道,天然化合物(Satoh等,2013)或通过破坏其负调节因子KEAP1激活NRF2导致髓样白细胞(Kong等,2011)和巨噬细胞(Lin等, 2008)的强效抗炎作用。 在观察性研究中,NFE2L2中的多态性与转录活性降低相关,与炎症性肠病(Arisawa等,2008b)和慢性胃炎(Arisawa等,2007)的风险增加相关。NRF2的免疫调节作用的一个例子是在中枢神经系统。受损神经元释放出趋化因子,一种特异性激活小胶质细胞中磷脂酰肌醇3激酶/ AKT(PI3K / AKT)途径的趋化因子,导致GSK-3β的抑制和NRF2的上调(LastresBecker等,2014)。在这项研究中,来自AD和进行性核上性麻痹患者的尸检显示出趋化因子水平的补偿性增加以及上调的NRF2蛋白,表明该途径有助于限制病变大脑中的炎症反应。
MS是一种慢性炎性疾病,其特征在于自身反应性免疫细胞浸润到中枢神经系统中。NRF2的缺失加剧了EAE的发展,EAE是MS的小鼠模型(Johnson等人,2010)。与NRF2缺乏相关的部分影响可能与HO-1水平降低有关。因此,具有骨髓特异性HO-1缺陷的小鼠表现出较高的病变发生率,伴随着抗原呈递细胞的活化和炎性Th17和髓鞘特异性T细胞的浸润(Tzima等人,2009)。敲除KEAP1(Kobayashi等,2016)或用各种激活NRF2的小分子治疗(Buendia等,2016)抑制了疾病的发展和严重程度。NRF2在活跃的MS病变中强烈上调,并且NRF2应答基因的表达主要在初始髓鞘破坏的区域中被发现(Licht-Mayer等,2015)。在MS脑中,NRF2及其靶标NADPH:醌氧化还原酶(NQO1)和HO-1主要在浸润性巨噬细胞中表达,在较小程度上在星形胶质细胞中表达,最有可能作为对病理性ROS形成的代偿性反应。相反,在少突胶质细胞中缺乏NRF2和抗氧化基因表达,这可能是它们在MS中的损伤和损失的基础(van Horssen等,2010)。由于免疫和氧化还原稳态的改变,MS患者外周血单核细胞中HO-1表达降低,并且在疾病恶化期间下调(Fagone等,2013)。值得注意的是,干扰素-β治疗患者的基因表达谱分析鉴定出NRF2是长期抗氧化反应和神经元保存的潜在介质(Croze等,2013)。
SLE由高氧化环境、失调的细胞死亡和去除死细胞的缺陷而突出,这导致细胞坏死作为自身抗原的来源。NRF2缺陷的雌性小鼠随年龄增长而发展为类似于SLE的多器官自身免疫性疾病,其特征为DNA氧化增加,脂质过氧化,脾细胞凋亡,抗双链DNA抗体和史密斯抗原的存在,以及伴随重要的组织损伤(血管炎,肾小球肾炎,肝炎和心肌炎)(Li等,2004)。只有雌性小鼠出现SLE进展的事实表明,雌性特异性因子可能有助于打破对自身抗原的免疫耐受(Li等,2004)。NRF2缺乏还导致CD4 + T细胞的增殖反应增强、CD4 + / CD8 +比率改变、以及SLE中促炎性Th17的促进(Ma等人,2006; Zhao等人,2016)。事实上,NRF2耗竭与狼疮性肾炎发展过程中的Th17分化和功能有关,这似乎是通过调节细胞因子信号传导抑制因子3 /磷酸化信号转导和转录激活因子(STAT)3途径和IL-1b信号来调节的(Zhao 等,2016)。此外,Nrf2-/ -小鼠的唾液腺显示出强烈的淋巴细胞浸润,使人想起Sjögren综合征,这通常与SLE有关(Ma等,2006)。SLE患者表现出氧化性DNA损伤修复机制的改变(Evans等,2000),高血清氧化蛋白水平,载脂蛋白C3(Morgan等,2007),氧化磷脂和抗氧化修饰脂蛋白的自身抗体(Frostegard等,2005)。NRF2多态性尚未与SLE易感性相关,尽管SNP rs35652124与墨西哥女性儿童期发病肾炎风险增加有关(Cordova等,2010)。
类风湿关节炎(RA)是一种全身炎症性疾病,具有复杂但仍难以捉摸的自身免疫特性,在炎症关节中,中性粒细胞、巨噬细胞和淋巴细胞被积极募集和激活。这导致促炎介质如ROS / RNS,类二十烷酸,细胞因子(IL-17,TNF-α,干扰素-?,IL-6和IL-1β)和分解代谢酶的分泌增加,这些促炎介质引发滑膜成纤维细胞的过度增殖, 关节肿胀,软骨和骨的逐渐破坏(Roberts等,2015)。NRF2基因的缺失增加了实验性RA模型中关节改变的易感性。例如,在表达T细胞受体KRN和主要组织相容性复合体II类分子A(g7) (K/BN关节炎模型)的小鼠中,以及在抗体诱导的关节炎中,NRF2缺乏加速了发病率并加重了疾病进程( Maicas等,2011; Wu等,2016b)。NRF2缺乏显着上调炎症细胞的迁移、环加氧酶-2和诱导型一氧化氮合酶的表达、ROS和RNS的产生以及促炎细胞因子和趋化因子的释放。此外,NRF2可能是关节炎中骨代谢的保护因子(Maicas等,2011),NRF2 / HO-1活化在RA动物模型和人滑膜成纤维细胞中发挥抗炎和抗氧化作用(Wu等,2016b)。有趣的是,抗风湿金(I)化合物通过激活NRF2和上调HO-1和GCLC来刺激抗氧化反应,证明其对RA的临床疗效(Kobayashi 等, 2016)。此外,NRF2 / HO-1激活介导了滑膜细胞凋亡的诱导和西洛他唑对促炎细胞因子产生的抑制(Park等,2010)以及H2S和相关化合物的抗炎作用,它们能通过对KEAP1的半胱氨酸残基进行硫化物修饰(Wu等,2016b)。其他诱导NRF2和HO-1信号传导的药物,如瑞巴派特,可以转导人和小鼠CD4+ T细胞向免疫抑制性Treg表型的分化,并通过特异性抑制STAT3来抑制TCD4+细胞向炎性Th17细胞的分化(Moon等,2014)。发炎的滑膜内过量的ROS产生似乎有助于RA的发病机制,因为患者的ROS形成,脂质过氧化,蛋白质氧化,DNA损伤和抗氧化防御机制活性降低显着增加,所有这些都促成了组织损伤和疾病进展(Datta等,2014)。响应于病理性ROS的形成,NRF2途径在RA患者的滑膜细胞和抗体诱导的关节炎小鼠的关节中被激活,但是该反应显然不足以抵消疾病进展(Wu等,2016b)。
白癜风是一种皮肤炎症性疾病,其特征在于表皮中ROS的积累,其参与黑素细胞的死亡。这些分子修饰DNA和黑素体蛋白,形成自身抗原并激活针对黑素细胞的自身免疫应答(Xie等,2016)。 遗传学研究发现NRF2启动子SNPs与白癜风易感性有关,如-650位点的SNP (Guan等, 2008),而rs35652124的C等位基因在汉族人群中被证明具有保护作用(Song等, 2016)。NRF2及其下游解毒靶基因NQO1,GCLC和GCLM在白癜风患者的表皮中上调,表明这种防御机制的激活不足(Natarajan等,2010)。
病理性ROS形成可能在慢性肺纤维化的发病机制中起作用。早期研究表明,博来霉素诱导的肺纤维化在Nrf2-/- 中比在野生型小鼠中更严重(Cho等,2004)。事实上,野生型小鼠通过上调NRF2诱导抗氧化和抗炎反应,而这在Nrf2-/-小鼠中无法实现。后来证实,患有特发性肺纤维化或慢性结节病/过敏性肺炎的患者表现出NRF2的表达增加,并且支气管肺泡灌洗液中的低摩尔.wt.抗氧化剂水平增加,如尿酸,抗坏血酸,视黄醇和α-生育酚 ,表明对ROS挑战的适应性反应不成功(Markart等,2009)。机制地,NRF2缺乏增加肌成纤维细胞分化,而用SFN药理诱导NRF2导致肌成纤维细胞数量减少和转化生长因子-β(TGF-β)的促纤维化作用减弱(Artaud-Macari等人,2013)。
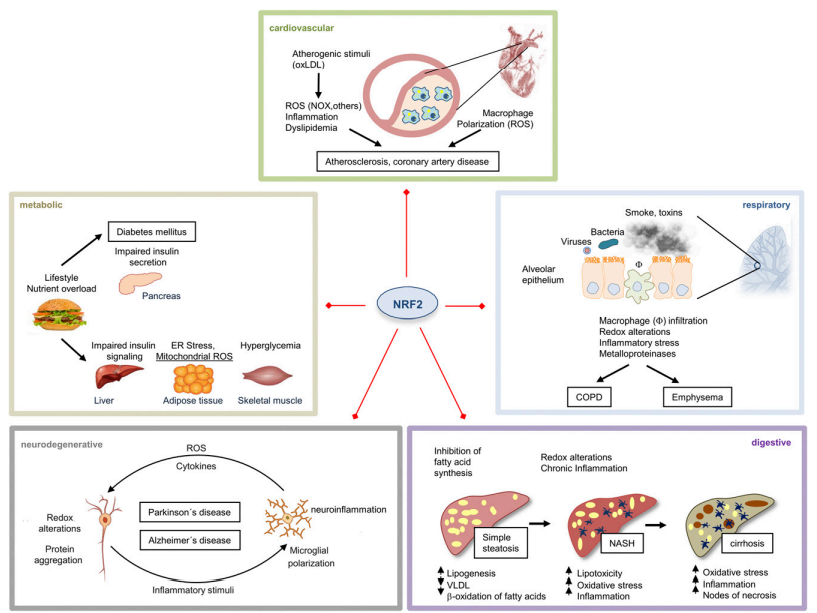
图8. NRF2在慢性病的常见机制和病理表型中的作用。该图提供了从图5的NRF2疾病组中提取的一些实例。这些疾病的常见病理机制包括异常高的ROS水平和参与组织损伤的低度慢性炎症。NRF2通过调节许多细胞保护基因的表达,提供针对这些和其他组织特异性改变的细胞保护特征。
GI的微生物群的共生作用之一是释放适量的ROS,其引起由NRF2介导的上皮细胞和浸润的免疫细胞中的细胞保护反应(Jones等,2015)。此外,在真核生物中受NRF2转录控制的细胞保护分子也可以由共生细菌产生。例如,微生物群中的HO-1同源物可能对胃肠道内稳态有很大的贡献,这可以用于治疗一氧化碳在肠道的局部递送(Onyiah等, 2014)。
NRF2在维持胃肠道稳态方面的作用已被证实,使NRF2这一转录因子成为IBD中有希望的治疗靶点。因此,几种化合物和膳食补充剂可能表现出有益效果,如褪黑激素,3-(3-吡啶基亚甲基)-2-吲哚满酮,丁酸盐,干酪乳杆菌,左旋肉碱,4-乙烯基-2,6-二甲氧基苯酚(卡诺尔), 乳枸杞(脱脂牛奶中枸杞的配方产品)等(Orena等,2015)。因此,明确NRF2在消化道慢性和急性疾病中的参与情况,对于更好地指导NRF2通路的调节治疗方法至关重要。
肝脏也是抵抗食物异生素的第一道防线。因此,NRF2疾病组强调该转录因子在与肝损伤相关的病理表型中的相关性也就不足为奇了。早期使用Nrf2-/-小鼠模型证明其对对乙酰氨基酚诱导的肝细胞损伤、苯并[a]芘诱导的肿瘤形成以及Fas-和TNF-α介导的肝细胞凋亡具有保护作用(Aleksunes和Manautou,2007)。Nrf2-/-小鼠对化学毒性的较高敏感性与解毒酶的基础表达和诱导表达降低相关。在人类中,导致NRF2表达降低的三种NRF2启动子SNPs的功能性单倍型与胃粘膜炎症的发展显著相关,无论是独立地还是通过与幽门螺杆菌感染相互作用(Arisawa等,2007)。对原发性胆汁性胆管炎患者NRF2转录特征的分析表明,这些患者表现出NRF2表达降低以及HO-1和GCLC蛋白水平低,这些损伤在肝硬化患者中更为严重(Wasik等,2017)。
病理性ROS形成是非酒精性脂肪性肝炎(NASH)患者肝细胞损伤和疾病进展的关键机制(图8)。该疾病分两个阶段发展,一个是肝细胞中脂肪酸的逐渐积累,另一个是肝损伤和炎性病理性ROS的形成(Wang等,2018)。因此,喂食高脂肪饮食(HFD)的小鼠出现了单纯的脂肪变性,其特征是肝脏脂肪沉积增加而没有炎症或纤维化,但Nrf2-/-小鼠出现加剧的肝脏脂肪变性和大量炎症,与NASH一致(Reccia等,2017)。然而有趣的是,肝细胞特异性KEAP1的缺失在减少肝脏脂肪变性的同时,并没有改变NASH发展过程中的炎症反应,提示其具有代偿机制(Ramadori等, 2016)。至少在NASH的大鼠模型中,饮食NRF2激活剂减弱了肝纤维化的进展(Shimozono等,2013)。在NASH患者的肝脏活组织检查中病理性ROS形成的标志物增加,并且NRF2特征增加,表明其试图减少氧化剂和炎症负荷(Takahashi等人,2014)。
NRF2在内皮稳态中最相关的靶点之一是HO-1,其通常与铁蛋白的上调相平行,因此降低游离铁水平,并阻止芬顿型反应。由HO-1和胆绿素还原酶的联合活性产生的胆红素是清除ROS / RNS的最强大的内源性抗氧化剂之一(Jansen等,2010),并且在体外预防脂质过氧化方面非常有效( Stocker等,1987)。Hmox1-/-小鼠显示出慢性缺氧引起的肺动脉高压升高(Christou等,2000),药理学HO-1诱导可改善糖尿病并发症(Kruger等,2006),以及硝酸甘油诱导的血管功能障碍( 硝酸盐耐受性)(Wenzel等,2007)。最近,Hmox1-/-小鼠在血管紧张素II响应中显示NADPH氧化酶-2上调、血管病理ROS形成、炎症标志物、内皮功能障碍和血压升高(Wenzel et al., 2015)。事实上,高血清胆红素水平与冠状动脉疾病的发病率呈负相关(Hopkins等,1996)。胆红素阻止血管NADPH氧化酶的活化(Kwak等,1991),其参与心血管疾病的发展(Griendling和FitzGerald,2003a,b; Harrison等,2003; Jay等,2006)。外周动脉疾病是动脉粥样硬化的常见表现,患有外周动脉疾病的患者HO-1水平降低(Signorelli等, 2016)。
NRF2缺乏对高血糖的影响首先在Nrf2-/-小鼠中显示,其中氧化和亚硝化改变增强并导致早期肾损伤(Yoh等,2008)。 在随后的研究中,链脲佐菌素诱导的糖尿病Nrf2-/-小鼠表现出加剧的肾小球损伤,同时ROS产生增加和促纤维化标志物TGF-β和纤连蛋白表达的增加(Jiang等人,2010)。在这个糖尿病模型中,NRF2对血视网膜屏障功能障碍和糖尿病视网膜病变的进展具有保护作用(Xu等, 2014)。同样,HFD诱导的血管ROS水平的增加在Nrf2-/-小鼠中显着加剧,并伴有严重的内皮功能障碍,表现为乙酰胆碱诱导的主动脉松弛减弱和细胞间粘附分子-1和TNF-α表达增加( Ungvari等,2011)。
NRF2在组织特异性胰岛素抵抗中起复杂作用。因此,与野生型小鼠相比,HFD喂养的Nrf2-/-小鼠由于肝脏和骨骼肌中胰岛素信号的增强而表现出更好的胰岛素敏感性,但相反,这些小鼠由于与病理ROS形成相关的肝脂毒性过高而产生严重的NASH (Meakin等, 2014)。因此,该研究将肝胰岛素抵抗与NASH的发展分离。根据这些数据,随后的研究表明,由于CYP2A5酶的表达减弱,HFD喂养的Nrf22-/-小鼠的肝脏由于GSH的显着消耗而表现出更高的病理性ROS形成(Cui等人,2013)。NRF2在肝细胞中的敲低增强了棕榈酸诱导的细胞凋亡,棕榈酸是一种在胰岛素抵抗性肥胖患者中高度升高的脂肪酸。这种效应与病理性ROS的产生增加相关,再次强化了NRF2在NASH进展中的关键作用(Pilar Valdecantos等,2015)。
为了进一步研究NRF2在代谢综合征中的作用,我们在瘦素缺乏(ob/ob)小鼠模型中去除NRF2,该模型具有极好的正能量平衡(Xue等, 2013)。有趣的是,全身ob / ob / Nrf2-/-小鼠或脂肪细胞特异性ob/ob/ Nrf2-/-小鼠显示白色脂肪量减少,显示NRF2是脂肪生成的关键参与者。这些小鼠具有更严重的代谢综合征,其特征在于高脂血症,加重的胰岛素抵抗和高血糖,表明代谢综合征与病理性ROS形成之间的机制联系。
另一组研究评估了持续诱导NRF2在葡萄糖代谢中的作用。NRF2基因诱导利用Keap1的一个亚型等位基因(Keap1flox/- 突变体)降低肥胖糖尿病db/db小鼠的血糖,是通过抑制肝细胞中cAMP-CREB信号传导以及其他糖异生基因(例如过氧化物酶体增殖激活受体辅激活因子-1α)来抑制肝葡萄糖6磷酸酶(Uruno等,2013)。此外,Keap11敲低小鼠中NRF2活性的增强增加了肝脏中AMP活化蛋白激酶(AMPK)的磷酸化,以及骨骼肌中的胰岛素信号传导,导致葡萄糖耐量的显着改善(Xu等,2013)。由于NRF2在T2DM背景下的多效活性,所有这些和其他研究的结果证明需要设计多种遗传和药理学策略来阐明参与全身葡萄糖稳态控制的组织中的全部NRF2功能。
除了年龄,体重和血糖等糖尿病因素外,与NRF2相关的遗传因素在人类中研究较少。在中国人群中,SNP rs6721961与病理性ROS形成和新诊断的T2DM风险有关,并且还可能导致胰岛素分泌能力受损和胰岛素抵抗增加(Wang等,2015)。在墨西哥混血男性中,相同的SNP与糖尿病相关(Jimenez-Osorio等,2017)。在一项汉族志愿者参与的病例对照研究中,发现T2DM患者无论有无并发症,包括周围神经病变、肾病、视网膜病变、足溃疡和微血管病变,NFE2L2基因4个SNPs的基因型和等位频率存在显著差异(Xu等, 2016b)。
Ramsey等人(2007)证明了NRF2在PD患者的多巴胺能神经元中的核定位。其他研究发现淀粉样蛋白前体蛋白和tau损伤神经元表达NRF2及其靶SQSTM1水平升高,可能是通过自噬清除这些毒性蛋白的补偿机制(Lastres-Becker等,2014; Pajares等,2016)。与这些结果一致,AD和PD脑中HO-1、NQO1、GCLM和SQSTM1水平升高(van Muiswinkel等, 2004;Cuadrado等,2009;Schipper等,2009;LastresBecker等,2016)。此外,与NRF2表达相关的细胞保护蛋白,如NQO1和SQSTM1,在路易体中被部分隔离,表明PD患者中NRF2特征的神经保护能力受损(Lastres-Becker等,2016)。这种差异的一种可能解释是NRF2及其靶基因水平可能在衰老和疾病进展过程中发生变化。
NFE2L2的一些SNP单倍型与ALS,AD或PD的风险降低或延迟发作相关。在关于先前与高基因表达相关的三种功能性启动子SNPs的两项研究中分析了ALS的发病。有趣的是,这种单倍型与ALS发病延迟4年有关(Bergstrom等,2014),但另一项研究未发现明确的相关性(LoGerfo等,2014)。关于AD,一个单倍型等位基因与AD的2年早期发病相关,表明NFE2L2基因的变体可能影响AD进展(von Otter等,2010b)。已经更详细地分析了NFE2L2与PD的遗传关联。NFE2L2启动子中的三个SNPs(rs6721961,rs6706649和rs35652124)在病例对照研究中被证明为保护性单倍型(von Otter等,2010a)。这种单倍型在瑞典队列中延迟了疾病的发作,甚至降低了波兰队列中PD的风险。这些结果得到了四项新的独立的欧洲病例对照研究的支持(von Otter等,2014),但未在台湾人群中复制(Chen等,2013),这表明种族和环境因素存在差异。另一种方法是将来自嗅黏膜的PD细胞暴露于烟提物或杀虫剂中,以评估基因-环境的相互作用,并且鉴定了几种影响对这些毒素的易感性的SNPs(Todorovic等,2015)。总而言之,NRF2的轻微激活(例如对于NFE2L2基因的某些功能性单倍型所发现的)应该足以触发大脑中的保护机制。
不断增加的ROS水平可以通过改变基因组稳定性来维持肿瘤发生,同时激活特定的氧化还原信号转导和促进肿瘤细胞存活和增殖的炎症过程(Reuter等,2010)。因此,NRF2的上调代表癌细胞适应的机制,以耐受推动肿瘤进展的高ROS水平(Schumacker,2006)以及维持导致肿瘤复发和远处转移形成的癌症干细胞(Ryoo等,2016)。例如,来自人类结肠直肠肿瘤的癌症干细胞中的NRF2特征指出了由高水平的GCLC,谷胱甘肽过氧化物酶和硫氧还蛋白还原酶-1介导的保护机制,这是这些细胞抵抗应激物和化学治疗剂的能力的基础(Emmink等,2013)。从这个角度来看,NRF2在癌细胞中表现得像一个致癌基因,通过诱导ARE介导的细胞保护反应的慢性激活,可以帮助其适应其氧化环境(Panieri和Santoro,2016)。已经报道了NRF2的恶性激活的几种机制,包括体细胞突变,表观遗传学和致癌信号传导改变。
据报道,沿着NFE2L2的编码序列在癌症中接近600个体细胞突变(Gao等人,2017)。在图9中,我们显示了来自10,000名癌症患者的数据集的结果(Zehir等人,2017)。在大多数情况下,这些突变改变了NRF2的DLG和ETGE基序与KEAP1的相互作用,从而诱导NRF2在几种实体瘤(包括食道癌,皮肤癌,肺癌和喉癌)中的过度活化(Kim等,2010b; Taguchi和 Yamamoto,2017)。例如,在晚期食管鳞状细胞癌中,NRF2功能获得性突变与肿瘤复发和预后不良相关,其原因包括增殖增加、独立依附生存以及对化疗和放疗的抵抗(Shibata 等, 2011)。KEAP1基因中的功能丧失突变在一些实体肿瘤例如肺癌中也是常见的(Singh等人,2006)。基于这一通路调控β-TrCP/NRF2的有力证据,奇怪的是在NRF2与β-TrCP的交界面未发现干扰体突变。这一事实表明,由于未知的原因,这些突变是不可生存的,或者β-TrCP逃逸导致NRF2水平的升高不足以驱动致癌性。
尽管如此,体细胞突变仅在一小部分癌症患者中引起长期/慢性NRF2激活。在基因表达水平上,有趣的是位于人NRF2基因的ARE增强子的SNP rs6721961(-617C>A)的等位基因消除了NRF2的自身诱导,这与这些癌症患者的显着存活相关(Okano等,2013)。已经在肺肿瘤中描述了由于KEAP1的三个CpG位点的启动子高甲基化引起的表观遗传变化,导致随后的NRF2活化,其可以通过5-氮杂-2’-脱氧胞苷处理逆转NRF2的活化(Wang等,2008)。最近已经回顾了miRNAs在NRF2水平的转录后调节中的作用(Kurinna和Werner,2015)。简而言之,miR200a靶向人乳腺癌细胞中的KEAP1 mRNA,导致其降解并随后激活NRF2(Eades等,2011)。反过来,miR28促进NRF2 mRNA的降解(Yang等人,2011)。
致癌基因或突变的肿瘤抑制因子可以增强癌症中NRF2的活化。KRAS,BRAF或c-MYC的内源性致癌等位基因上调NRF2,可能是通过致癌基因介导的ROS和随后的KEAP1慢性失活(DeNicola等,2011)。肿瘤抑制因子p53的突变形式,通过增强营养摄取和构建块的合成来维持癌细胞的生长,也可以通过与结合NRF2启动子的Sp1转录因子的串扰来上调NRF2(Tung等,2015)。
各种蛋白激酶对NRF2(磷酸-NRF2)的磷酸化是107种肝细胞癌中潜在的激活机制。增加的磷酸化NRF2水平与表现出这种独特表型的患者的KEAP1表达降低和5年总体存活率差有关(Chen等,2016)。此外,磷酸酶和张力蛋白同源物(PTEN)肿瘤抑制因子的突变维持过度活跃的和致癌的磷脂酰肌醇-3-激酶(PI3K)-AKT信号传导,并由于为NRF2蛋白酶体降解的PTEN / GSK-3 / β-TrCP途径的下调而导致NRF2活性增加。(Rada等,2011,2012; Cuadrado,2015)。针对PTEN / GSK-3 / β-TrCP途径的治疗干预措施应考虑到GSK-3既可作为肿瘤抑制因子又可作为肿瘤启动子,并且还与癌症干细胞的产生有关(McCubrey等,2014)。
一些应激诱导的蛋白质与KEAP1相互作用,因此在癌细胞中,这些蛋白与KEAP1结合的NRF2竞争。因此,NRF2逃脱了KEAP1介导的降解。NRF2与KEAP1结合的竞争对手之一是自噬衔接蛋白SQSTM1的磷酸化形式,该蛋白发生在癌细胞用于维持自身生长的选择性自噬过程中(Shimizu等, 2016)。还证明了促进癌症干细胞中细胞周期停滞的细胞周期蛋白依赖性激酶抑制剂p21通过其KRR基序与NRF2中的DLG和ETGE基序的相互作用抑制NRF2与KEAP1的结合(Chen等人,2009)。最近,显示具有ETGE基序的二肽基肽酶3可以与NRF2竞争结合KEAP1(Hast等人,2013)。可能由氧化还原状态的长期改变诱导的二肽基肽酶3的过表达与ARE基因的表达增加和预后不良相关,特别是在雌激素受体阳性乳腺癌中(Lu等人,2017)。
NRF2诱导导致癌症进展的代谢变化。例如,一项多平台非靶向代谢组学研究确定了乳腺肿瘤样本中代谢物变化的模式(Tang等,2014)。他们发现GSH和3-(4-羟基苯基)乳酸通过与NRF2的相互作用与BRCA1参与氧化还原稳态呈正相关。代谢组学研究还表明,NRF2可以增加癌细胞中的有氧糖酵解,以支持其高能量需求。这通过NRF2介导的锰-超氧化物歧化酶表达的诱导发生,导致线粒体过氧化氢的产生升高和AMPK的活化。该过程由caveolin-1调节,其直接结合NRF2和KEAP1,并阻碍NRF2活化并因此阻碍糖酵解转变。这显然解释了为什么通常更具侵略性的糖酵解肿瘤具有caveolin-1低/ Mn超氧化物歧化酶高的表型(Hart等,2016)。NRF2还可以驱动葡萄糖和谷氨酰胺向肿瘤细胞增殖所需的合成代谢途径(Mitsuishi等,2012)。在PI3K-AKT信号通路活跃和KEAP1活性缺失的情况下,NRF2被证明可以诱导癌症中葡萄糖代谢从糖酵解向合成代谢途径(嘌呤合成)的转移(Mitsuishi等, 2012;Xu等, 2016a)。以上这一点被强调通过NRF2介导的参与磷酸戊糖途径的基因转录和NADPH(葡萄糖6磷酸脱氢酶;磷酸葡萄糖酸脱氢酶,苹果酸酶1,异柠檬酸脱氢酶1,转酮醇酶和转醛酶)的产生以及涉及嘌呤核苷酸合成(磷酸核糖焦磷酸酰胺转移酶,亚甲基四氢叶酸脱氢酶2)的相关基因。
NRF2的激活赋予癌细胞生长优势的事实可能会争辩说,在这项工作中提到的慢性疾病NRF2的药理学活化可能意味着患癌症的高风险。然而,必须考虑到NRF2的致癌活性需要其基因或KEAP1发生突变,从而导致NRF2信号的持续高水平诱导。但在药理学治疗中并非如此,在药理学治疗中,调节药物剂量和NRF2活性是可能的。此外,经验证据表明,参与NRF2激活剂临床试验的受试者没有显示出更高的癌症风险。自从2013年获得监管机构批准以来,已经服用NRF2活化剂富马酸二甲酯数年的MS患者就是最好的例证。相反,在癌症患者中使用NRF2抑制剂可能导致NRF2疾病组中描述的病理表型的表现。当NRF2抑制剂到达临床时,这种可能性需要进一步研究。
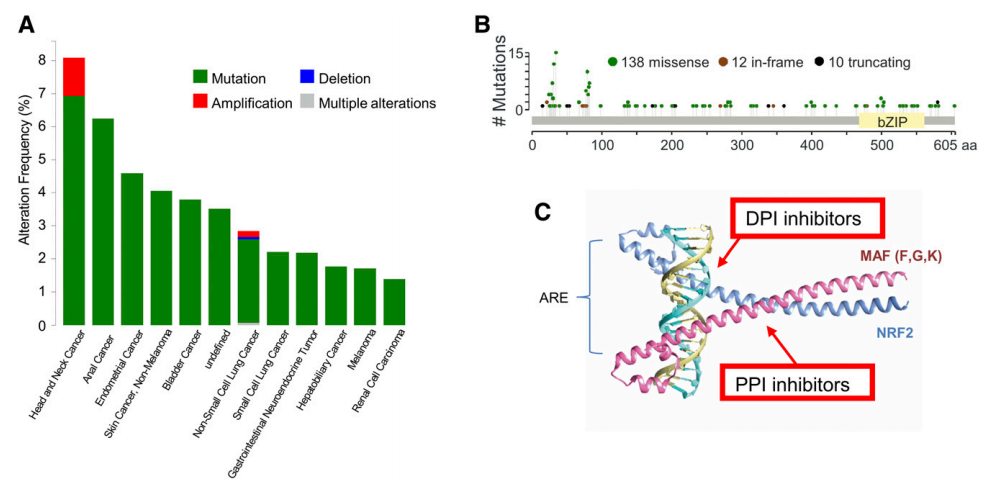
图9. 在MSK IMPACT临床测序组(MSKCC)研究(Zehir等,2017)的肿瘤中发现的体细胞突变和抑制NRF2的药理学策略。(A)具有NRF2突变的肿瘤的百分比。 (B)沿NRF2多肽的突变分布。(C)NRM2 / MAFF异二聚体与ARE元件之间相互作用的PyMOL表示。 蓝色:NRF2; 粉红色:MAFF。 红色箭头表示可能通过小分子抑制NRF2的机制,这些小分子可以靶向NRF2和MAF蛋白之间相互作用的bZip结构域(PPI抑制剂)或NRF2-MAF异二聚体与ARE相互作用的界面[DNA-蛋白质相互作用(DPI)]
NRF2调制剂的至少30项近期专利在世界国际产权组织中被收录。这些专利正在保护查尔酮衍生物,新型酰胺三萜衍生物,氘取代的富马酸衍生物,3-烷基氨基-1H-吲哚丙烯酸酯衍生物,甲醇酰胺,含有活化乙烯基的苄基衍生物,穿心莲内酯或[S]+阿扑吗啡,以及倍半萜内酯衍生物(Sun等,2017)。尽管从临床前概念验证的角度来看,大多数这些化合物在某种程度上被证明是有用的,但它们的临床价值迄今通常非常有限。其中只有少数药物进入了临床试验,而食品和药物管理局(Food and Drug Administration)或欧洲药品管理局(European Medicines Agency)等监管机构批准的药物更少。我们在这项研究中讨论了转化管道中最发达的NRF2激活剂。
富马酸酯是KEAP1改性剂的最突出的例子,并且富马酸二甲酯(DMF)是迄今为止唯一的食品和药物管理局和欧洲药品管理局批准的药物,其注册为NRF2活化剂。单酯形式的DMF,富马酸单甲酯(MMF)被描述为其活性代谢物。DMF和MMF是迈克尔受体,其直接与KEAP1中存在的半胱氨酸残基反应(Lin等,2011)。
从NRF2的功能尚不清楚开始,DMF和其他富马酸酯已经用于治疗牛皮癣超过50年。该化合物在欧洲以商品名Fumaderm获得许可。临床试验显示,在DMF治疗12-16周后,银屑病面积和严重程度指数降低至50%-80%(Altmeyer等,1994; Mrowietz等,1998)。最近,在III期试验(BRIDGE)中,DMF已证明其在治疗患有中度至重度慢性斑块状银屑病的成人中的功效(Mrowietz等,2017)。富马酸盐在银屑病病变缓解中的作用机制包括外周T细胞数量的减少以及从Th1向Th2免疫应答的转变(Ghoreschi等,2011; Tahvili等,2015)。在另一种自身免疫性疾病SLE中,富马酸酯已被用作治疗严重、广泛和难治性皮肤表现的全身联合疗法(Saracino和Orteu,2017)。
2013年,DMF以商品名Tecfidera被批准用于治疗MS(Xu等,2015)。在MS患者中使用DMF是通过在EAE的MS小鼠模型中获得的阳性结果推动的。对疾病进程和组织学的显着治疗效果与脊髓中巨噬细胞介导的炎症显着减少有关。血液中的多重细胞因子分析证明DMF处理的动物中抗炎细胞因子IL-10的增加(Schilling等人,2006)。此外,DMF还改善了野生型中髓鞘,轴突和神经元的保存,但在Nrf2-/-小鼠中没有(Ellrichmann等,2011)。在人类中,DMF显示MS病变和年复发率显着降低(Schimrigk等,2006)。两项III期临床试验DEFINE和CONFIRM证实了这些结果(Fox等,2012; Gold等,2012)。因此,DMF目前被用作复发缓解型MS的第一线治疗,其不能通过传统疗法治疗。新的DMF配方正在测试和申请专利,以提高药物的生物利用度和功效(Sun等,2017)。例如,MMF已被用于开发第二代NRF2诱导物作为前药(Zeidan等,2014)。铅化合物ALKS-8700,一种MMF的2-(2,5-二氧代-1-吡咯烷基)乙酯衍生物,在体内迅速转化为MMF,因此提高了其生物利用度并减少了胃肠道副作用。ALKS-8700目前正在进行III期临床试验(EVOLVE MS)。
DMF和MMF调节免疫应答。例如,它们通过减少炎性细胞因子的释放并因此减少DCs处理抗原的能力来抑制DCs的成熟。此外,DMF和MMF激活天然杀伤细胞以裂解DCs并增强DCs和T细胞的凋亡(Ghoreschi等,2011; Al-Jaderi和Maghazachi,2015)。因此,DMF和MMF阻碍T细胞介导的自身反应。一些研究表明,DMF还通过触发GSH耗竭诱导II型DCs,这导致HO-1活性增强和STAT1磷酸化抑制。这些经典的II型DCs抑制Th1和Th17介导的反应,支持Th2的反应。此外,通过DCs增加IL-10的产生有利于CD4+T细胞向抑制性Treg表型的分化(Ockenfels等,1998; Ghoreschi等,2011)。DMF还抑制NF-κB的核转位(Peng等,2012),从而在小胶质细胞和星形胶质细胞(Brennan等,2017)以及外周血单核细胞(Eminel等,2017)中产生炎症介质,如TNF-α,IL-1β,IL-6,趋化因子,粘附分子和一氧化氮。此外,DMF发挥抗血管生成作用,其依赖于内皮细胞中血管内皮生长因子受体-2表达的下调(Meissner等,2011)。最近的研究结果表明,DMF减少了CD4+,CD8+,Th1和Th17细胞的数量,而CD4+/CD8+比例和Th2亚群在这些患者的血液中增加。有趣的是,DMF/MMF对T细胞活化的抑制作用主要局限于记忆T细胞(Wu等, 2017)。DMF或MMF的这些免疫调节活性对于保护少突胶质细胞免受ROS诱导的细胞毒性具有重要意义(Scannevin等,2012)。
其他机制可以解释NF-κB的抑制,而与NRF2活化无关。因此,DMF可能与几种调节NF-kB信号传导的蛋白质中的半胱氨酸残基相互作用(Blewett等,2016)。此外,DMF可以抑制泛素偶联酶,从而防止NF-κB的IkB抑制因子在IL-1β或Toll样受体激动剂的作用下降解(McGuire等, 2016)。此外,DMF直接结合蛋白激酶C-θ中的特定半胱氨酸残基,这是参与T细胞受体信号传导的关键激酶(Blewett等,2016)。此外,MMF和DMF激活羟基羧酸受体-2,导致NF-κB的抑制和促炎细胞因子和粘附分子的下调(Chen等,2014; Gillard等,2015)并导致中性粒细胞浸润减少 (Chen等,2014)。尽管这些NRF2非依赖性作用与EAE的急性炎症期相关,但DMF在慢性自身免疫性脱髓鞘中的神经保护功效取决于NRF2活化(Linker等,2011)。在Nrf2-/-和野生型小鼠中DMF治疗的临床益处与炎性Th1和Th17细胞的减少以及抗炎M2单核细胞的诱导相关。同时,在野生型中观察到CD80和CD86共刺激分子的表达降低,但在Nrf2-/-小鼠中未观察到,表明至少这些作用是NRF2依赖性的(Schulze-Topphoff等,2016)。
DMF用于治疗自身免疫疾病的成功表明,具有由慢性,低级炎症和病理性ROS形成强调的共同病理机制的其他疾病可能受益于该药物的重新定位。在亨廷顿舞蹈病的小鼠模型中,DMF治疗保留了存活率、肌肉功能和体重,这与完整神经元数量的增加有关(Ellrichmann等,2011)。此外,在最近的PD临床前研究中,使用该疾病的α-突触核蛋白病模型,由于自噬诱导受损,DMF在野生型中具有神经保护作用,但在Nrf2-/-小鼠中则不具有神经保护作用(Lastres-Becker等,2016)。
DMF被证明可预防糖尿病小鼠的内皮功能障碍和心血管病理性ROS形成和炎症(Sharma等,2017),并且在注射链脲佐菌素后载脂蛋白E缺陷小鼠中动脉粥样硬化、肾功能不全和其他糖尿病并发症减少(Tan等,2014)。此外,一些研究表明,DMF可能通过抑制NF-κB途径发挥抗肿瘤活性,因此在治疗侵袭性癌症中增加了治疗价值(Kastrati等,2016)。DMF是网络药理学方法中老药新用概念的相关实例。
合成三萜类化合物是2-氰基-3,12-二氧代 - 油酸-1,9(11) - 二烯-28-酸盐(CDDO; 巴多索隆,RTA401)的衍生物,其类似于天然产物齐墩果酸。它们通过其α-β不饱和支架表现出迈克尔受体活性,并且代表了最有效的NRF2诱导剂(Sun等,2017)。它们与KEAP1的C151相互作用并阻碍其与CUL3的相互作用,从而导致NRF2激活(Cleasby等,2014)。原理证明研究强烈支持合成的三萜类化合物用于退行性疾病,并且正在成为Reata / Abbott作为炎症抗氧化剂调节剂的深入研究的焦点。例如,CDDO-咪唑(CDDO-Im RTA403)在野生型而非Nrf2-/-小鼠的腹膜中性粒细胞中诱导各种抗氧化基因(Hmox1,Gclc,Gclm和Nqo1)的表达,并减弱LPS诱导的ROS产生和产生和促炎细胞因子的产生,从而降低死亡率(Thimmulappa等,2006b)。CDDO-乙基酰胺(RTA405)和CDDO-CDDO-三氟乙基酰胺(RTA 404)在毒素诱导的PD模型(1-甲基-4-苯基-1,2,3,6-四氢吡啶)中测量的所有终点均具有显着作用 (Kaidery等,2013)。在MS的EAE模型中,CDDOCDDO-三氟乙基酰胺抑制炎症、病理性ROS形成和髓鞘变性(Pareek等,2011)。
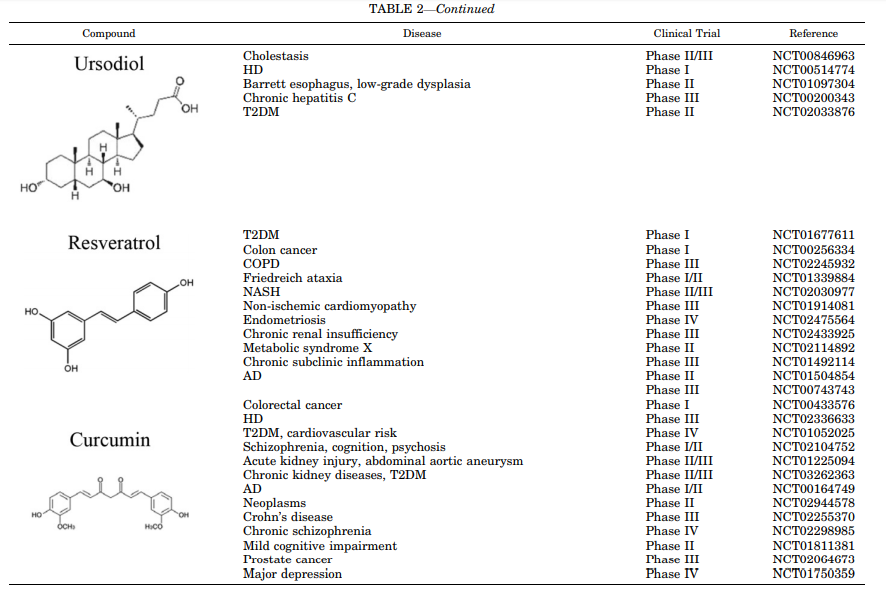
CDDO-甲酯(CDDO-Me,RTA 402)是在用于治疗糖尿病肾病的临床试验中达到的第一个CDDO(Pergola等,2011)。虽然第二阶段的结果非常令人鼓舞,但由于心血管安全问题(Zhang,2013),CDDO-Me后来在第三阶段(BEACON试验)被撤回,这与NRF2无关,但最有可能是脱靶改变内皮素信号传导(de Zeeuw等,2013; Chin等,2014)。目前,CDDO-Me正在进行临床研究,作为Alport综合征和肺动脉高压的潜在治疗方法(表2)。为了改善其安全性,进一步的研究导致了CDDO-二氟丙酰胺(RTA-408, omaveloxone)的开发,该药物目前正在进行二期试验,用于治疗弗里德雷希的共济失调、眼部炎症和眼部手术后疼痛。
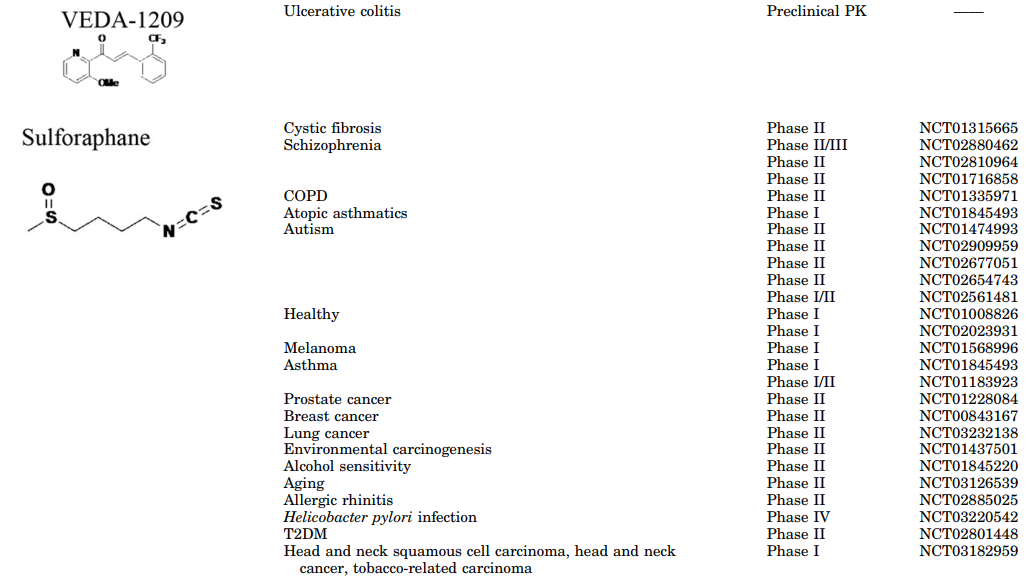
奥替普拉是一种有机硫化合物,用作抗血吸虫病药物,目前正在进行III期试验,用于治疗非酒精性脂肪性肝炎。用于治疗亨廷顿氏病的高级临床试验正在开发中,米诺环素是一种抗生素,由于NRF2活化而具有神经保护作用(Kuang等,2009)。用于治疗急性肾病的I期临床研究中的另一种NRF2诱导剂是CXA-10,一种通过激活NRF2具有抗炎特性的硝基脂肪酸(Batthyany和Lopez,2015)。近年来,许多具有相同作用机制的NRF2诱导剂已被描述(Buendia等, 2015a,b, 2016),其中一些还处于临床前研究阶段,如化合物VEDA-1209,它是一种查尔酮衍生物,具有良好的抗炎作用,可用于溃疡性结肠炎的治疗。
SFN是由有机硫化合物萝卜硫苷的酶促裂解产生的异硫氰酸酯,其存在于西兰花、卷心菜和其他十字花科植物的芽苗中。催化反应由植物中发现的黑芥子酶和胃肠道的微生物群驱动(Kensler等,2013)。催化反应由植物中发现的黑芥子酶和胃肠道的微生物群驱动(Kensler等,2013)。最近,SFN已经通过化学合成获得(Kim等,2015)。通过向患有T2DM的患者施用含有SFN的花椰菜芽粉,实现了SFN向临床的转化(Bahadoran等,2012)。西兰花粉降低血浆丙二醛和氧化低密度脂蛋白(LDL),提高总抗氧化能力。心血管危险因素如血清甘油三酯,氧化LDL/LDL比率和血浆致动脉粥样硬化指数(甘油三酯对数/高密度脂蛋白比率)也降低。此外,促炎标志物如C-反应蛋白和IL-6减少。在最近的一项研究中,SFN作为浓缩西兰花芽提取物使用,通过NRF2的核转位抑制肝细胞葡萄糖的生成,并降低参与糖异生的关键酶的表达。此外,SFN降低了患有T2DM的肥胖患者的空腹血糖和糖化血红蛋白(Axelsson等,2017)。SFN诱导的NRF2活化通过降低ROS负荷和抑制NF-κB和TGF-β1信号传导途径来保护肾细胞免于狼疮性肾炎(Jiang等,2014a)。
关于神经退行性疾病,已经表明SFN穿过血脑屏障并提供足够的脑生物利用度来激活NRF2特征并减少LPS诱发的神经炎症,这反映在促炎性标志物(诱导型一氧化氮合酶、IL-6、TNF-α)的减少中和海马中的小神经胶质细胞增生(Innamorato等,2008)。SFN还保护多巴胺能神经元免受帕金森病毒1-甲基-4-苯基-1,2,3,6-四氢吡啶的影响,并减弱星形胶质细胞增生和小神经胶质细胞增生(Jazwa等,2011)。与这些发现一致,SFN降低了磷酸化tau水平并增加了Beclin-1和LC3-II,表明NRF2活化可能通过大脑中的自噬促进这种毒性蛋白的降解(Jo等,2014)。SFN处理的脊髓损伤大鼠炎症因子水平明显降低,挫伤体积减小,协调性改善(Wang等, 2012)。该药还通过保留血脑屏障和减少病理性ROS形成和炎症细胞数量来改善EAE(Li等,2013)。SFN迄今已用于至少32项针对慢性疾病的临床研究,如癌症、哮喘、慢性肾病、T2DM、囊性纤维化、自闭症和精神分裂症(Duran等,2016; Houghton等,2016)(表2)。
表2. 选择的NRF2诱导剂作为KEAP1的亲电子修饰剂
该引用对应于ClinicalTrials.gov中的代码。




总之,这些观察结果为其他SFN衍生化合物的开发铺平了道路,这些化合物表现出更好的药代动力学特征。SFN是一种在亲水性介质中稳定性较差的油性物质。其物理化学特征促使Evgen Pharma(Wilmslow,Cheshire,England)开发出一种环糊精复合物制剂Sulforadex,该制剂正在进行用于治疗蛛网膜下腔出血的II期临床试验。SFN还与褪黑激素杂交以产生ITH12674,该化合物被设计为具有用于治疗脑缺血的双重药物-前药作用机制(Egea等人,2015)。
姜黄素是姜黄中发现的主要类姜黄素,已被用于治疗肥胖症、代谢综合征和前驱糖尿病。采用气相色谱-电子冲击质谱法对姜黄素对大鼠肝脏的影响进行了非靶向代谢组学研究。间断摄入姜黄素可上调NRF2,并在抗肝损伤方面发挥抗氧化和抗炎作用(Qiu等,2016)。口服姜黄素可有效降低血清甘油三酯、IL-1β、IL-4和血管内皮生长因子,以及增加血液中脂联素水平。在T2DM患者中,姜黄素降低空腹血糖,糖化血红蛋白,血清游离脂肪酸,甘油三酯和尿酸的水平,并增加脂蛋白脂肪酶的水平(Na等,2013; Chuengsamarn等,2014)。
白藜芦醇是一种保护植物免受真菌感染的多酚,存在于葡萄皮、红葡萄酒、浆果和许多其他植物中。白藜芦醇通过下调KEAP1表达和激活蛋白去乙酰化酶sirtuin-1来激活NRF2信号通路,从而发挥抗氧化作用(Ungvari等, 2010)。在健康受试者中,白藜芦醇的饮食施用防止了血浆中胆固醇、内毒素、促氧化剂和炎性标记物(p47phox,KEAP1,IL-1β和TNF-α)的升高。这些事件与NRF2活性的升高相关,NRF2活性升高是由NRF2靶标NQO1和谷胱甘肽S-转移酶的表达增强所确定的(Ghanim等,2011)。在T2DM患者中,治疗4周后胰岛素敏感性得到改善,这通过AKT增强胰岛素信号传导,病理性ROS形成减少和糖化血红蛋白水平降低来确定(Brasnyo等,2011; Bhatt等,2012)。总的来说,白藜芦醇在动物模型和患者中可预防高血压、高胆固醇血症、动脉粥样硬化、缺血性心脏病、糖尿病和代谢综合征等主要心血管、炎症、氧化和代谢并发症(Xia等, 2017)。
经常被忽视的问题是亲电KEAP1抑制剂缺乏选择性。亲电试剂与细胞中不同的亲核试剂发生反应,从而表现出非靶向和非预期的副作用。例如,CDDO-Im可与500多种不同的目标相互作用(Yore等,2011)。通常,几种蛋白磷酸酶在其催化中心含有对氧化还原敏感的半胱氨酸,并且一些KEAP1抑制剂可以修饰和灭活这些磷酸酶,因此扰乱信号传导网络。这些磷酸酶之一是PTEN(Lee等,2002; Kitagishi和Matsuda,2013; Han等,2015)。PTEN的催化C124残基可以通过与强亲电试剂例如CDDO-Im(Pitha-Rowe等,2009)和叔丁基氢醌(Rojo等,2014b)形成加合物来修饰。然后,PI3K/AKT通路的激活增加涉及GSK-3的抑制和NRF2随后的稳定(图2C) (Rada等, 2011, 2012)。此外,KEAP1与其他同样含有高亲和力结合基序ETGE的蛋白相互作用(Hast等, 2013),如Bcl-2和IKKβ (Kim等, 2010a;Cazanave等,2014)。因此,从KEAP1缺陷细胞获得的一些结果可能不一定与NRF2活化有关。
最近对KEAP1/NRF2相互作用的新多肽和小分子抑制剂的发现进行了综述(Abed等, 2015;Jiang等, 2016)。简而言之,最初使用表面等离振子共振和荧光偏振测定评估一系列截短的NRF2肽作为PPI的直接抑制剂(Hu等,2013)。抑制能力最小的肽序列为LDE-ETGE-FL的9-mer序列(Chen等, 2011;Inoyama等,2012)。与此同时,Wells和合作者(Hancock等,2013)使用噬菌体展示文库结合高通量荧光偏振测定法搜索新推定的肽配体。他们发现,与单独的天然肽相比,基于NRF2和SQSTM1的ETGE基序的杂合肽对KEAP1具有更高的结合活性。为了促进细胞摄取,设计一种肽,其中ETGE基序与HIV-Tat蛋白的细胞转导结构域和钙蛋白酶的切割序列(DEETGE-Cal-Tat)融合。该肽在脑缺血的小鼠模型中显示出神经保护和认知保护作用(Tu等,2015)。
已经描述了五个PPI抑制剂家族:四氢异喹啉(Jnoff等,2014; Richardson等,2015),硫嘧啶(Marcotte等,2013),萘(Jiang等,2014b),咔唑酮(Ranjan等,2014)和尿素衍生物(Sato等,2013)。表3汇总了最近针对这些小分子的专利。 尽管这些化合物非常有前途,但仍需要证明它们对KEAP1/NRF2相互作用具有选择性,因为KEAP1也至少靶向Bcl2和IKK(Kim等,2010a; Hast等,2013; Cazanave等,2014)。
从可用文库中索引的大量化合物中,化合物LH601,苯磺酰基嘧啶酮2,N-苯基-苯磺酰胺和一系列1,4-二苯基-1,2,3-三唑可能是KEAP1抑制PPI非常适合的候选者(Hu等,2013; Jnoff等,2014; Bertrand等,2015; Wen等,2015; Nasiri等,2016)。这些研究详细描述了与KEAP1的原子相互作用、亲和力和结合的热力学参数。这些化合物的治疗功效将在未来的工作中进行分析,其中应该解决安全性,效力和血脑屏障渗透性。
表3. 选择的NRF2诱导剂充当NRF2-KEAP1蛋白质 - 蛋白质相互作用抑制剂



从概念上讲,β-TrCP-磷酸化NRF2相互作用的抑制剂也应该导致NRF2活化,因为它们应该破坏NRF2降解的这一分支(图2)。β-TrCP的β-螺旋和含有NRF2磷酸二核的肽之间的分子相互作用已经通过NMR解决(Rada等,2012)。正如KEAP1/EGTE所发生的那样,最相关的氨基酸似乎是β-TrCP的几个精氨酸残基,其与DpSGIpS基序的两个磷酸丝氨酸相互作用。然而,能够抑制β-TrCP-磷酸化NRF2相互作用的小分子的发现仍在进行中。
已经开发了其他策略来抑制NRF2抑制因子BACH1,这是一种bZip蛋白,可与MAF蛋白形成异二聚体并阻断ARE基因的表达。已经在体外描述了HPP-4382化合物对BACH1的有效抑制(Attucks等,2014),但是,在完整的临床试验之前,必须在体内证明HPP-4382的安全性和功效特征。考虑到其他途径也可能影响NRF2的活性,我们有理由推测,组合方法将是激活该转录因子的最佳途径。
天然产物如鸦胆子苦醇(Ren等,2011; Olayanju等,2015),赭曲霉毒素A(Tarumoto等,2004; Limonciel和Jennings,2014)和葫芦巴碱(Arlt等,2013)也有被发现抑制NRF2。然而,他们的作用机制还没有完全明白。事实上,与现有化合物相关的一个重要问题是它们可能具有的深远的脱靶效应。例如,最近发现鸦胆子苦醇对蛋白质合成具有普遍和非特异性抑制作用,导致NRF2水平下降,而且许多其他快速转换蛋白下降,因此最近不鼓励使用鸦胆子苦醇(Harder et al。,2017))。类似地,兽医实践中使用的抗原虫剂卤虫酮通过抑制NRF2积累来增强癌细胞的化学敏感性,但这种效应似乎是间接的,它通过抑制核糖基转移RNA的合成,这是NRF2以及许多其他含脯氨酸的蛋白质的核糖体转化所强烈需要的(Tsuchida等,2017)。
最近报道了一种识别选择性NRF2抑制剂的新方法,即使用小分子抑制剂的高通量定量筛选(Singh等, 2016)。作者确定了一种名为ML385的一流化合物,它最有可能阻止NRF2与其他bZip共激活因子的结合。该化合物阻断NRF2转录活性并使KEAP1缺陷细胞对卡铂和其他化学治疗药物敏感。需要进一步的研究来确认ML385是否对NRF2具有选择性,或者它是否也抑制其他bZip转录因子。
鉴于NRF2在多种肿瘤病理表型中具有良好的全身效应,用小分子抑制剂特异性靶向NRF2似乎提供了一种良好的临床途径。然而,有必要确定用NRF2抑制剂治疗癌症是否会增加NRF2疾病组中的其他病理表型的风险。
二甲双胍是T2DM的一线单药治疗。根据图6,它为与葡萄糖代谢相关的病理表型的NRF2亚群提供治疗益处。事实上,SFN可降低肝脏葡萄糖的产生,并改善T2DM患者的血糖控制(Axelsson等,2017)。有趣的是,一些证据表明,二甲双胍可能有效预防NRF2疾病组中的其他非糖尿病病理表型,包括心血管疾病(Nesti和Natali,2017),呼吸系统疾病(Sato等,2016),消化系统(Bauer和Duca,2016),神经退行性疾病(Markowicz-Piasecka等,2017),自身免疫(Schuiveling等,2017)和肿瘤(Heckman-Stoddard等,2017)疾病。二甲双胍的作用机制尚不完全清楚,但它涉及抑制线粒体复合物I,从而增加AMP/ATP比值(El-Mir等,2000; Owen等,2000)并导致能量传感器AMPK的激活(Hardie,2004; Rena等,2017)。重要的是,AMPK激活NRF2(Wang等,2017a; Zhao等,2017),并且该轴的药理学靶向可减轻中风后的炎症(Wang等,2017c)或内毒素暴露(Ci等,2017; Lv等,2017)。事实上,二甲双胍以AMPK依赖的方式激活NRF2,从而抑制临床前啮齿动物短暂性全脑缺血模型的炎症反应(Ashabi等, 2015;Kaisar等,2017)。葡萄糖代谢和炎症可能不是二甲双胍/NRF2作用的唯一病理机制。实际上,已经描述了氧化还原的其他有益效果(Kocer等,2014; Kelleni等,2015)和蛋白质稳态(Tsai等,2017)。
他汀类药物可预防和减少心血管病变表型。除了降脂作用外,他汀类药物似乎可以预防与NRF2网络相关的病理机制,如炎症(Pantan等,2016; Wu等,2016a; Hwang等,2017)和病理性ROS形成( Abdanipour等,2014)。它们是3-羟基-3-甲基-戊二酰-CoA还原酶的竞争性抑制剂,其催化胆固醇合成中的限速反应。其他多效性影响包括转录因子Krüppel样因子2的上调,其在肝硬化进展期间早期诱导,并减轻肝血管功能障碍的发展(Marrone等,2015)。最近的证据表明,至少一些他汀类药物会激活NRF2。在分离的肝细胞中进行的一项研究中,高浓度的辛伐他汀激活了NRF2,可能作为一种防御机制(Cho等,2013)。用洛伐他汀预处理神经干细胞激活NRF2途径并引发针对过氧化氢诱导的细胞死亡的保护(Abdanipour等,2014)。在肝硬化中,辛伐他汀激活由Krüppel样因子2和NRF2形成的轴,以减少星状细胞的氧化负荷和炎症反应,改善肝纤维化,内皮功能障碍和门静脉高压。辛伐他汀激活NRF2的机制尚不完全清楚,但似乎涉及NRF2相互作用组中发现的元素,如丝裂原活化蛋白激酶、PI3K/AKT途径(Jang等,2016)和GSK-3(Lin等,2016)。
从图4的NRF2相互作用组可以推断出药物新用的其他病例,特别是信号激酶。如图2C所示,GSK-3磷酸化NRF2的Neh6结构域,导致β-TrCP的识别和进一步泛素依赖性蛋白酶体降解。GSK-3在没有刺激的情况下是活跃的,而在激活AKT和其他激酶的信号级联导致GSK-3在其N端伪底物域磷酸化时是不活跃的。因此,已知靶向信号激酶的药物可用于上调(GSK-3抑制剂)或下调(PI3K/AKT抑制剂)NRF2特征。
GSK-3参与NRF2疾病组中发现的至少一些病理表型,例如糖尿病和神经退行性疾病(Beurel等,2015; Maqbool和Hoda,2017)。从天然和合成来源发现了广泛的GSK-3抑制剂(Khan等,2017),但将GSK-3抑制剂重新利用来增加NRF2活性的最佳证据可能源于临床使用锂作为情绪稳定剂(Chiu等,2013)。虽然此时在NRF2疾病组中未发现双相情感障碍和抑郁,但很明显它们表现出神经炎症和退行性病变表型,至少在小鼠模型中暗示NRF2失调(Martin-de-Saavedra等,2013; Freitas等,2016; Yao等,2016)。
NRF2相互作用组也为阻断信号激酶从而激活GSK-3的癌症药物抑制NRF2提供了一个理由。例如,表皮生长因子受体抑制剂厄洛替尼导致NRF2抑制,参与非小细胞肺癌中的肿瘤细胞感觉(Xiaobo等,2016)。用于治疗肝细胞癌的激酶级联抑制剂索拉非尼也导致NRF2及其下游靶标金属硫蛋白-1(Houessinon等,2016)和亚甲基四氢叶酸脱氢酶1的抑制(Lee等人,2017)。
最后,在两项相关研究中,迄今为止一直在寻找可能影响NRF2调节的再利用药物。使用基于荧光相关光谱的筛选系统,1633种药物中的两种显着增加HepG2细胞中的NRF2蛋白水平:叶绿酸和bonaphton(Yoshizaki等,2017)。在另一项研究中,分析了连接图数据库,其包含用1309种试剂处理的人细胞系的基因表达谱(Lamb等,2006; Iorio等,2010)(Zhang等,2017),通过激活NRF2以寻找有潜力的氧化还原调节剂(Xiong等,2014)。该研究发现阿司咪唑是一种有效的抗组胺药,用于过敏性疾病,是一种新型的NRF2激活剂。
与NRF2转录相关的变化可用作监测药物功效的生物标志物,所述药物旨在通过黄嘌呤氧化酶和NADPH氧化酶抑制剂减少病理性ROS形成。同样,可以通过定义全局蛋白和基因表达谱来检测和监测环境化学品的暴露情况(Ghezzi等,2017a)。这种方法类似于第一阶段药物代谢酶的使用,其中细胞色素P450是各种异种生物通过Ah受体诱导的,可以作为海洋污染的指标(Cajaraville等, 2000)。12周内每日口服富马酸酯与银屑病患者皮肤中NRF2靶基因表达增加有关(Onderdijk等,2014)。同样,在接受每日剂量CDDO-Me治疗3周的癌症患者外周血单核细胞中,NQO1的mRNA水平增加了5倍(Hong等, 2012)。
使用NRF2的转录特征作为生物标记需要很好地了解ARE基因活化所涉及的机制,因为大多数NRF2靶标受另外的转录因子调节。因此,分析几种ARE基因的表达很重要。例如,一项使用NRF2作为肺鳞癌治疗反应预测因子的研究提出使用28个基因来定义NRF2激活谱(Cescon等, 2015)。
Abed DA, Goldstein M, Albanyan H, Jin H, and Hu L (2015) Discovery of direct inhibitors of Keap1-Nrf2 protein-protein interaction as potential therapeutic and preventive agents. Acta Pharm Sin B 5:285–299.
Aleksunes LM, Goedken M, and Manautou JE (2006) Up-regulation of NAD(P)H quinone oxidoreductase 1 during human liver injury. World J Gastroenterol 12: 1937–1940.
Aleksunes LM and Manautou JE (2007) Emerging role of Nrf2 in protecting against hepatic and gastrointestinal disease. Toxicol Pathol 35:459–473.
Al-Huseini LM, Aw Yeang HX, Sethu S, Alhumeed N, Hamdam JM, Tingle Y, Djouhri L, Kitteringham N, Park BK, Goldring CE, et al. (2013) Nuclear factorerythroid 2 (NF-E2) p45-related factor-2 (Nrf2) modulates dendritic cell immune function through regulation of p38 MAPK-cAMP-responsive element binding protein/activating transcription factor 1 signaling. J Biol Chem 288:22281–22288.
Al-Jaderi Z and Maghazachi AA (2015) Vitamin D3 and monomethyl fumarate enhance natural killer cell lysis of dendritic cells and ameliorate the clinical score in mice suffering from experimental autoimmune encephalomyelitis. Toxins (Basel) 7: 4730–4744.
Altmeyer PJ, Matthes U, Pawlak F, Hoffmann K, Frosch PJ, Ruppert P, Wassilew SW, Horn T, Kreysel HW, Lutz G, et al. (1994) Antipsoriatic effect of fumaric acid derivatives: results of a multicenter double-blind study in 100 patients. J Am Acad Dermatol 30:977–981.
Archer E, Pavela G, and Lavie CJ (2015) The inadmissibility of what we eat in America and NHANES dietary data in nutrition and obesity research and the scientific formulation of national dietary guidelines. Mayo Clin Proc 90:911–926.
Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, Fujita H, Hasegawa S, Takagi T, Wang FY, et al. (2007) The relationship between Helicobacter pylori infection and promoter polymorphism of the Nrf2 gene in chronic gastritis. Int J Mol Med 19:143–148.
Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, Fujita H, Yoshioka D, Okubo M, Hirata I, et al. (2008a) Nrf2 gene promoter polymorphism and gastric carcinogenesis. Hepatogastroenterology 55:750–754.
Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, Fujita H, Yoshioka D, Okubo M, Sakata M, et al. (2008b) Nrf2 gene promoter polymorphism is associated with ulcerative colitis in a Japanese population. Hepatogastroenterology 55:394–397.
Arlt A, Sebens S, Krebs S, Geismann C, Grossmann M, Kruse ML, Schreiber S, and Schäfer H (2013) Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 32: 4825–4835.
Artaud-Macari E, Goven D, Brayer S, Hamimi A, Besnard V, Marchal-Somme J, Ali ZE, Crestani B, Kerdine-Römer S, Boutten A, et al. (2013) Nuclear factor erythroid 2-related factor 2 nuclear translocation induces myofibroblastic dedifferentiation in idiopathic pulmonary fibrosis. Antioxid Redox Signal 18:66–79.
Ashabi G, Khalaj L, Khodagholi F, Goudarzvand M, and Sarkaki A (2015) Pretreatment with metformin activates Nrf2 antioxidant pathways and inhibits inflammatory responses through induction of AMPK after transient global cerebral ischemia. Metab Brain Dis 30:747–754.
Attucks OC, Jasmer KJ, Hannink M, Kassis J, Zhong Z, Gupta S, Victory SF, Guzel M, Polisetti DR, Andrews R, et al. (2014) Induction of heme oxygenase I (HMOX1) by HPP-4382: a novel modulator of Bach1 activity. PLoS One 9:e101044.
Aviello G and Knaus UG (2017) ROS in gastrointestinal inflammation: rescue or sabotage? Br J Pharmacol 174:1704–1718.
Axelsson AS, Tubbs E, Mecham B, Chacko S, Nenonen HA, Tang Y, Fahey JW, Derry JMJ, Wollheim CB, Wierup N, et al. (2017) Sulforaphane reduces hepatic glucose production and improves glucose control in patients with type 2 diabetes. Sci Transl Med 9:1–12.
Bahadoran Z, Mirmiran P, Hosseinpanah F, Rajab A, Asghari G, and Azizi F (2012) Broccoli sprouts powder could improve serum triglyceride and oxidized LDL/LDLcholesterol ratio in type 2 diabetic patients: a randomized double-blind placebocontrolled clinical trial. Diabetes Res Clin Pract 96:348–354.
Baillie JK, Arner E, Daub C, De Hoon M, Itoh M, Kawaji H, Lassmann T, Carninci P, Forrest AR, Hayashizaki Y, et al.; FANTOM Consortium (2017) Analysis of the human monocyte-derived macrophage transcriptome and response to lipopolysaccharide provides new insights into genetic aetiology of inflammatory bowel disease. PLoS Genet 13:e1006641.
Baird L and Dinkova-Kostova AT (2013) Diffusion dynamics of the Keap1-Cullin3 interaction in single live cells. Biochem Biophys Res Commun 433:58–65. Banning A and Brigelius-Flohé R (2005) NF-kappaB, Nrf2, and HO-1 interplay in redox-regulated VCAM-1 expression. Antioxid Redox Signal 7:889–899.
Barabási AL, Gulbahce N, and Loscalzo J (2011) Network medicine: a network-based approach to human disease. Nat Rev Genet 12:56–68.
Batthyany CI and Lopez GV (2015) Nitroalkene Tocopherols and Analogs Thereof for Use in the Treatment and Prevention of Inflammation Related Conditions, Complexa, Radnor, PA. Bauer PV and Duca FA (2016) Targeting the gastrointestinal tract to treat type 2 diabetes. J Endocrinol 230:R95–R113.
Bergström P, von Otter M, Nilsson S, Nilsson AC, Nilsson M, Andersen PM, Hammarsten O, and Zetterberg H (2014) Association of NFE2L2 and KEAP1 haplotypes with amyotrophic lateral sclerosis. Amyotroph Lateral Scler Frontotemporal Degener 15:130–137.
Bertrand HC, Schaap M, Baird L, Georgakopoulos ND, Fowkes A, Thiollier C, Kachi H, Dinkova-Kostova AT, and Wells G (2015) Design, synthesis, and evaluation of triazole derivatives that induce Nrf2 dependent gene products and inhibit the Keap1-Nrf2 protein-protein interaction. J Med Chem 58:7186–7194.
Beurel E, Grieco SF, and Jope RS (2015) Glycogen synthase kinase-3 (GSK3): regulation, actions, and diseases. Pharmacol Ther 148:114–131. Bhatt JK, Thomas S, and Nanjan MJ (2012) Resveratrol supplementation improves glycemic control in type 2 diabetes mellitus. Nutr Res 32:537–541.
Blewett MM, Xie J, Zaro BW, Backus KM, Altman A, Teijaro JR, and Cravatt BF (2016) Chemical proteomic map of dimethyl fumarate-sensitive cysteines in primary human T cells. Sci Signal 9:rs10.
Bolaños JP (2016) Bioenergetics and redox adaptations of astrocytes to neuronal activity. J Neurochem 139 (Suppl 2):115–125.
Bourdonnay E, Morzadec C, Fardel O, and Vernhet L (2009) Redox-sensitive regulation of gene expression in human primary macrophages exposed to inorganic arsenic. J Cell Biochem 107:537–547.
Bowie A and O’Neill LA (2000) Oxidative stress and nuclear factor-kappaB activation: a reassessment of the evidence in the light of recent discoveries. Biochem Pharmacol 59:13–23.
Brasnyó P, Molnár GA, Mohás M, Markó L, Laczy B, Cseh J, Mikolás E, Szijártó IA, Mérei A, Halmai R, et al. (2011) Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br J Nutr 106:383–389.
Brennan MS, Matos MF, Richter KE, Li B, and Scannevin RH (2017) The NRF2 transcriptional target, OSGIN1, contributes to monomethyl fumarate-mediated cytoprotection in human astrocytes. Sci Rep 7:42054.
Brüne B, Dehne N, Grossmann N, Jung M, Namgaladze D, Schmid T, von Knethen A, and Weigert A (2013) Redox control of inflammation in macrophages. Antioxid Redox Signal 19:595–637.
Buendia I, Gómez-Rangel V, González-Lafuente L, Parada E, León R, Gameiro I, Michalska P, Laudon M, Egea J, and López MG (2015a) Neuroprotective mechanism of the novel melatonin derivative Neu-P11 in brain ischemia related models. Neuropharmacology 99:187–195.
Buendia I, Michalska P, Navarro E, Gameiro I, Egea J, and León R (2016) Nrf2-ARE pathway: an emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases. Pharmacol Ther 157:84–104.
Buendia I, Navarro E, Michalska P, Gameiro I, Egea J, Abril S, López A, GonzálezLafuente L, López MG, and León R (2015b) New melatonin-cinnamate hybrids as multi-target drugs for neurodegenerative diseases: Nrf2-induction, antioxidant effect and neuroprotection. Future Med Chem 7:1961–1969.
Cajaraville MP, Bebianno MJ, Blasco J, Porte C, Sarasquete C, and Viarengo A (2000) The use of biomarkers to assess the impact of pollution in coastal environments of the Iberian Peninsula: a practical approach. Sci Total Environ 247: 295–311.
Cazanave SC, Wang X, Zhou H, Rahmani M, Grant S, Durrant DE, Klaassen CD, Yamamoto M, and Sanyal AJ (2014) Degradation of Keap1 activates BH3-only proteins Bim and PUMA during hepatocyte lipoapoptosis. Cell Death Differ 21: 1303–1312.
Cescon DW, She D, Sakashita S, Zhu CQ, Pintilie M, Shepherd FA, and Tsao MS (2015) NRF2 pathway activation and adjuvant chemotherapy benefit in lung squamous cell carcinoma. Clin Cancer Res 21:2499–2505.
Chen H, Assmann JC, Krenz A, Rahman M, Grimm M, Karsten CM, Köhl J, Offermanns S, Wettschureck N, and Schwaninger M (2014) Hydroxycarboxylic acid receptor 2 mediates dimethyl fumarate’s protective effect in EAE. J Clin Invest 124:2188–2192.
Chen J, Yu Y, Ji T, Ma R, Chen M, Li G, Li F, Ding Q, Kang Q, Huang D, et al. (2016) Clinical implication of Keap1 and phosphorylated Nrf2 expression in hepatocellular carcinoma. Cancer Med 5:2678–2687.
Chen W, Sun Z, Wang XJ, Jiang T, Huang Z, Fang D, and Zhang DD (2009) Direct interaction between Nrf2 and p21(Cip1/WAF1) upregulates the Nrf2-mediated antioxidant response. Mol Cell 34:663–673.
Chen Y, Inoyama D, Kong AN, Beamer LJ, and Hu L (2011) Kinetic analyses of Keap1-Nrf2 interaction and determination of the minimal Nrf2 peptide sequence required for Keap1 binding using surface plasmon resonance.
Chem Biol Drug Des 78:1014–1021. Chen YC, Wu YR, Wu YC, Lee-Chen GJ, and Chen CM (2013) Genetic analysis of NFE2L2 promoter variation in Taiwanese Parkinson’s disease. Parkinsonism Relat Disord 19:247–250.
Chin MP, Reisman SA, Bakris GL, O’Grady M, Linde PG, McCullough PA, Packham D, Vaziri ND, Ward KW, Warnock DG, et al. (2014) Mechanisms contributing to adverse cardiovascular events in patients with type 2 diabetes mellitus and stage 4 chronic kidney disease treated with bardoxolone methyl. Am J Nephrol 39: 499–508.
Chiu CT, Wang Z, Hunsberger JG, and Chuang DM (2013) Therapeutic potential of mood stabilizers lithium and valproic acid: beyond bipolar disorder. Pharmacol Rev 65:105–142.
Cho HY and Kleeberger SR (2010) Nrf2 protects against airway disorders. Toxicol Appl Pharmacol 244:43–56.
Cho HY, Marzec J, and Kleeberger SR (2015) Functional polymorphisms in Nrf2: implications for human disease. Free Radic Biol Med 88 (Pt B):362–372.
Cho HY, Reddy SP, Yamamoto M, and Kleeberger SR (2004) The transcription factor NRF2 protects against pulmonary fibrosis. FASEB J 18:1258–1260.
Cho YE, Moon PG, Lee JE, Singh TS, Kang W, Lee HC, Lee MH, Kim SH, and Baek MC (2013) Integrative analysis of proteomic and transcriptomic data for identification of pathways related to simvastatin-induced hepatotoxicity. Proteomics 13: 1257–1275.
Chowdhry S, Zhang Y, McMahon M, Sutherland C, Cuadrado A, and Hayes JD (2013) Nrf2 is controlled by two distinct b-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 32:3765–3781.
Christou H, Morita T, Hsieh CM, Koike H, Arkonac B, Perrella MA, and Kourembanas S (2000) Prevention of hypoxia-induced pulmonary hypertension by enhancement of endogenous heme oxygenase-1 in the rat. Circ Res 86: 1224–1229.
Chuengsamarn S, Rattanamongkolgul S, Phonrat B, Tungtrongchitr R, and Jirawatnotai S (2014) Reduction of atherogenic risk in patients with type 2 diabetes by curcuminoid extract: a randomized controlled trial. J Nutr Biochem 25:144–150.
Ci X, Zhou J, Lv H, Yu Q, Peng L, and Hua S (2017) Betulin exhibits antiinflammatory activity in LPS-stimulated macrophages and endotoxin-shocked mice through an AMPK/AKT/Nrf2-dependent mechanism. Cell Death Dis 8:e2798.
Cleasby A, Yon J, Day PJ, Richardson C, Tickle IJ, Williams PA, Callahan JF, Carr R, Concha N, Kerns JK, et al. (2014) Structure of the BTB domain of Keap1 and its interaction with the triterpenoid antagonist CDDO. PLoS One 9:e98896.
Comandini A, Marzano V, Curradi G, Federici G, Urbani A, and Saltini C (2010) Markers of anti-oxidant response in tobacco smoke exposed subjects: a data-mining review. Pulm Pharmacol Ther 23:482–492.
Córdova EJ, Velázquez-Cruz R, Centeno F, Baca V, and Orozco L (2010) The NRF2 gene variant, -653G/A, is associated with nephritis in childhood-onset systemic lupus erythematosus. Lupus 19:1237–1242.
Croze E, Yamaguchi KD, Knappertz V, Reder AT, and Salamon H (2013) Interferonbeta-1b-induced short- and long-term signatures of treatment activity in multiple sclerosis. Pharmacogenomics J 13:443–451.
Cuadrado A (2015) Structural and functional characterization of Nrf2 degradation by glycogen synthase kinase 3/b-TrCP. Free Radic Biol Med 88 (Pt B):147–157.
Cuadrado A, Martín-Moldes Z, Ye J, and Lastres-Becker I (2014) Transcription factors NRF2 and NF-kB are coordinated effectors of the Rho family, GTP-binding protein RAC1 during inflammation. J Biol Chem 289:15244–15258.
Cuadrado A, Moreno-Murciano P, and Pedraza-Chaverri J (2009) The transcription factor Nrf2 as a new therapeutic target in Parkinson’s disease. Expert Opin Ther Targets 13:319–329.
Cui Y, Wang Q, Li X, and Zhang X (2013) Experimental nonalcoholic fatty liver disease in mice leads to cytochrome p450 2a5 upregulation through nuclear factor erythroid 2-like 2 translocation. Redox Biol 1:433–440.
Datta S, Kundu S, Ghosh P, De S, Ghosh A, and Chatterjee M (2014) Correlation of oxidant status with oxidative tissue damage in patients with rheumatoid arthritis. Clin Rheumatol 33:1557–1564.
de la Vega MR, Dodson M, Gross C, Manzour H, Lantz RC, Chapman E, Wang T, Black SM, Garcia JG, and Zhang DD (2016) Role of Nrf2 and autophagy in acute lung injury. Curr Pharmacol Rep 2:91–101.
DeNicola GM, Karreth FA, Humpton TJ, Gopinathan A, Wei C, Frese K, Mangal D, Yu KH, Yeo CJ, Calhoun ES, et al. (2011) Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 475:106–109.
de Zeeuw D, Akizawa T, Audhya P, Bakris GL, Chin M, Christ-Schmidt H, Goldsberry A, Houser M, Krauth M, Lambers Heerspink HJ, et al.; BEACON Trial Investigators (2013) Bardoxolone methyl in type 2 diabetes and stage 4 chronic kidney disease. N Engl J Med 369:2492–2503.
Diotallevi M, Checconi P, Palamara AT, Celestino I, Coppo L, Holmgren A, Abbas K, Peyrot F, Mengozzi M, and Ghezzi P (2017) Glutathione fine-tunes the innate immune response toward antiviral pathways in a macrophage cell line independently of its antioxidant properties. Front Immunol 8:1239.
Duran CG, Burbank AJ, Mills KH, Duckworth HR, Aleman MM, Kesic MJ, Peden DB, Pan Y, Zhou H, and Hernandez ML (2016) A proof-of-concept clinical study examining the NRF2 activator sulforaphane against neutrophilic airway inflammation. Respir Res 17:89.
Eades G, Yang M, Yao Y, Zhang Y, and Zhou Q (2011) miR-200a regulates Nrf2 activation by targeting Keap1 mRNA in breast cancer cells. J Biol Chem 286: 40725–40733.
Egea J, Buendia I, Parada E, Navarro E, Rada P, Cuadrado A, López MG, García AG, and León R (2015) Melatonin-sulforaphane hybrid ITH12674 induces neuroprotection in oxidative stress conditions by a ‘drug-prodrug’ mechanism of action. Br J Pharmacol 172:1807–1821.
Ellrichmann G, Petrasch-Parwez E, Lee DH, Reick C, Arning L, Saft C, Gold R, and Linker RA (2011) Efficacy of fumaric acid esters in the R6/2 and YAC128 models of Huntington’s disease. PLoS One 6:e16172.
El-Mir MY, Nogueira V, Fontaine E, Avéret N, Rigoulet M, and Leverve X (2000) Dimethylbiguanide inhibits cell respiration via an indirect effect targeted on the respiratory chain complex I. J Biol Chem 275:223–228.
Eminel S, Jin N, Rostami M, Dibbert S, Mrowietz U, and Suhrkamp I (2017) Dimethyl- and monomethylfumarate regulate indoleamine 2,3-dioxygenase (IDO) activity in human immune cells. Exp Dermatol 26:685–690.
Emmink BL, Verheem A, Van Houdt WJ, Steller EJ, Govaert KM, Pham TV, Piersma SR, Borel Rinkes IH, Jimenez CR, and Kranenburg O (2013) The secretome of colon cancer stem cells contains drug-metabolizing enzymes. J Proteomics 91:84–96.
Evans MD, Cooke MS, Akil M, Samanta A, and Lunec J (2000) Aberrant processing of oxidative DNA damage in systemic lupus erythematosus. Biochem Biophys Res Commun 273:894–898.
Fagone P, Patti F, Mangano K, Mammana S, Coco M, Touil-Boukoffa C, Chikovani T, Di Marco R, and Nicoletti F (2013) Heme oxygenase-1 expression in peripheral blood mononuclear cells correlates with disease activity in multiple sclerosis. J Neuroimmunol 261:82–86.
Finkelstein R, Fraser RS, Ghezzo H, and Cosio MG (1995) Alveolar inflammation and its relation to emphysema in smokers. Am J Respir Crit Care Med 152:1666–1672.
Fox RJ, Miller DH, Phillips JT, Hutchinson M, Havrdova E, Kita M, Yang M, Raghupathi K, Novas M, Sweetser MT, et al.; CONFIRM Study Investigators (2012) Placebo-controlled phase 3 study of oral BG-12 or glatiramer in multiple sclerosis. N Engl J Med 367:1087–1097.
Freitas AE, Egea J, Buendia I, Gómez-Rangel V, Parada E, Navarro E, Casas AI, Wojnicz A, Ortiz JA, Cuadrado A, et al. (2016) Agmatine, by improving neuroplasticity markers and inducing Nrf2, prevents corticosterone-induced depressivelike behavior in mice. Mol Neurobiol 53:3030–3045.
Frijhoff J, Winyard PG, Zarkovic N, Davies SS, Stocker R, Cheng D, Knight AR, Taylor EL, Oettrich J, Ruskovska T, et al. (2015) Clinical relevance of biomarkers of oxidative stress. Antioxid Redox Signal 23:1144–1170.
Frostegård J, Svenungsson E, Wu R, Gunnarsson I, Lundberg IE, Klareskog L, Hörkkö S, and Witztum JL (2005) Lipid peroxidation is enhanced in patients with systemic lupus erythematosus and is associated with arterial and renal disease manifestations. Arthritis Rheum 52:192–200.
Gao J, Chang MT, Johnsen HC, Gao SP, Sylvester BE, Sumer SO, Zhang H, Solit DB, Taylor BS, Schultz N, et al. (2017) 3D clusters of somatic mutations in cancer reveal numerous rare mutations as functional targets. Genome Med 9:4.
Ghanim H, Sia CL, Korzeniewski K, Lohano T, Abuaysheh S, Marumganti A, Chaudhuri A, and Dandona P (2011) A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, highcarbohydrate meal. J Clin Endocrinol Metab 96:1409–1414.
Ghezzi P, Floridi L, Boraschi D, Cuadrado A, Manda G, Levic S, D’Acquisto F, Hamilton A, Athersuch TJ, and Selley L (2017a) Oxidative stress and inflammation induced by environmental and psychological stressors: a biomarker perspective. Antioxid Redox Signal [published ahead of print].
Ghezzi P, Jaquet V, Marcucci F, and Schmidt HHHW (2017b) The oxidative stress theory of disease: levels of evidence and epistemological aspects. Br J Pharmacol 174:1784–1796.
Ghoreschi K, Brück J, Kellerer C, Deng C, Peng H, Rothfuss O, Hussain RZ, Gocke AR, Respa A, Glocova I, et al. (2011) Fumarates improve psoriasis and multiple sclerosis by inducing type II dendritic cells. J Exp Med 208:2291–2303.
Gillard GO, Collette B, Anderson J, Chao J, Scannevin RH, Huss DJ, and Fontenot JD (2015) DMF, but not other fumarates, inhibits NF-kB activity in vitro in an Nrf2-independent manner. J Neuroimmunol 283:74–85.
Goh KI, Cusick ME, Valle D, Childs B, Vidal M, and Barabási AL (2007) The human disease network. Proc Natl Acad Sci USA 104:8685–8690.
Gold R, Kappos L, Arnold DL, Bar-Or A, Giovannoni G, Selmaj K, Tornatore C, Sweetser MT, Yang M, Sheikh SI, et al.; DEFINE Study Investigators (2012) Placebo-controlled phase 3 study of oral BG-12 for relapsing multiple sclerosis. N Engl J Med 367:1098–1107.
Gonzalez-Donquiles C, Alonso-Molero J, Fernandez-Villa T, Vilorio-Marqués L, Molina AJ, and Martín V (2017) The NRF2 transcription factor plays a dual role in colorectal cancer: a systematic review. PLoS One 12:e0177549.
Goven D, Boutten A, Leçon-Malas V, Marchal-Sommé J, Amara N, Crestani B, Fournier M, Lesèche G, Soler P, Boczkowski J, et al. (2008) Altered Nrf2/Keap1- Bach1 equilibrium in pulmonary emphysema. Thorax 63:916–924.
Griendling KK and FitzGerald GA (2003a) Oxidative stress and cardiovascular injury: part I: basic mechanisms and in vivo monitoring of ROS. Circulation 108: 1912–1916.
Griendling KK and FitzGerald GA (2003b) Oxidative stress and cardiovascular injury: part II: animal and human studies. Circulation 108:2034–2040.
Guan CP, Zhou MN, Xu AE, Kang KF, Liu JF, Wei XD, Li YW, Zhao DK, and Hong WS (2008) The susceptibility to vitiligo is associated with NF-E2-related factor2 (Nrf2) gene polymorphisms: a study on Chinese Han population. Exp Dermatol 17: 1059–1062.
Guney E, Menche J, Vidal M, and Barábasi AL (2016) Network-based in silico drug efficacy screening. Nat Commun 7:10331.
Guney E and Oliva B (2014) Analysis of the robustness of network-based diseasegene prioritization methods reveals redundancy in the human interactome and functional diversity of disease-genes. PLoS One 9:e94686.
Han SJ, Ahn Y, Park I, Zhang Y, Kim I, Kim HW, Ku CS, Chay KO, Yang SY, Ahn BW, et al. (2015) Assay of the redox state of the tumor suppressor PTEN by mobility shift. Methods 77–78:58–62.
Hancock R, Schaap M, Pfister H, and Wells G (2013) Peptide inhibitors of the Keap1- Nrf2 protein-protein interaction with improved binding and cellular activity. Org Biomol Chem 11:3553–3557.
Harder B, Tian W, La Clair JJ, Tan AC, Ooi A, Chapman E, and Zhang DD (2017) Brusatol overcomes chemoresistance through inhibition of protein translation. Mol Carcinog 56:1493–1500.
Hardie DG (2004) The AMP-activated protein kinase pathway–new players upstream and downstream. J Cell Sci 117:5479–5487.
Harrison D, Griendling KK, Landmesser U, Hornig B, and Drexler H (2003) Role of oxidative stress in atherosclerosis. Am J Cardiol 91:7A–11A.
Hart PC, Ratti BA, Mao M, Ansenberger-Fricano K, Shajahan-Haq AN, Tyner AL, Minshall RD, and Bonini MG (2016) Caveolin-1 regulates cancer cell metabolism via scavenging Nrf2 and suppressing MnSOD-driven glycolysis. Oncotarget 7: 308–322.
Harvey CJ, Thimmulappa RK, Sethi S, Kong X, Yarmus L, Brown RH, Feller-Kopman D, Wise R, and Biswal S (2011) Targeting Nrf2 signaling improves bacterial clearance by alveolar macrophages in patients with COPD and in a mouse model. Sci Transl Med 3:78ra32.
Hast BE, Goldfarb D, Mulvaney KM, Hast MA, Siesser PF, Yan F, Hayes DN, and Major MB (2013) Proteomic analysis of ubiquitin ligase KEAP1 reveals associated proteins that inhibit NRF2 ubiquitination. Cancer Res 73:2199–2210.
Hayes JD and Dinkova-Kostova AT (2014) The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem Sci 39: 199–218.
Heckman-Stoddard BM, DeCensi A, Sahasrabuddhe VV, and Ford LG (2017) Repurposing metformin for the prevention of cancer and cancer recurrence. Diabetologia 60:1639–1647.
Hidalgo CA, Blumm N, Barabási AL, and Christakis NA (2009) A dynamic network approach for the study of human phenotypes. PLOS Comput Biol 5:e1000353.
Hong DS, Kurzrock R, Supko JG, He X, Naing A, Wheler J, Lawrence D, Eder JP, Meyer CJ, Ferguson DA, et al. (2012) A phase I first-in-human trial of bardoxolone methyl in patients with advanced solid tumors and lymphomas. Clin Cancer Res 18:3396–3406.
Hopkins PN, Wu LL, Hunt SC, James BC, Vincent GM, and Williams RR (1996) Higher serum bilirubin is associated with decreased risk for early familial coronary artery disease. Arterioscler Thromb Vasc Biol 16:250–255.
Houessinon A, François C, Sauzay C, Louandre C, Mongelard G, Godin C, Bodeau S, Takahashi S, Saidak Z, Gutierrez L, et al. (2016) Metallothionein-1 as a biomarker of altered redox metabolism in hepatocellular carcinoma cells exposed to sorafenib. Mol Cancer 15:38.
Houghton CA, Fassett RG, and Coombes JS (2016) Sulforaphane and other nutrigenomic Nrf2 activators: can the clinician’s expectation be matched by the reality? Oxid Med Cell Longev 2016:7857186.
Hu L, Magesh S, Chen L, Wang L, Lewis TA, Chen Y, Khodier C, Inoyama D, Beamer LJ, Emge TJ, et al. (2013) Discovery of a small-molecule inhibitor and cellular probe of Keap1-Nrf2 protein-protein interaction. Bioorg Med Chem Lett 23: 3039–3043.
Hua CC, Chang LC, Tseng JC, Chu CM, Liu YC, and Shieh WB (2010) Functional haplotypes in the promoter region of transcription factor Nrf2 in chronic obstructive pulmonary disease. Dis Markers 28:185–193.
Hur W and Gray NS (2011) Small molecule modulators of antioxidant response pathway. Curr Opin Chem Biol 15:162–173.
Hwang AR, Han JH, Lim JH, Kang YJ, and Woo CH (2017) Fluvastatin inhibits AGE-induced cell proliferation and migration via an ERK5-dependent Nrf2 pathway in vascular smooth muscle cells. PLoS One 12:e0178278.
Innamorato NG, Rojo AI, García-Yagüe AJ, Yamamoto M, de Ceballos ML, and Cuadrado A (2008) The transcription factor Nrf2 is a therapeutic target against brain inflammation. J Immunol 181:680–689.
Inoyama D, Chen Y, Huang X, Beamer LJ, Kong AN, and Hu L (2012) Optimization of fluorescently labeled Nrf2 peptide probes and the development of a fluorescence polarization assay for the discovery of inhibitors of Keap1-Nrf2 interaction. J Biomol Screen 17:435–447.
Iorio F, Isacchi A, di Bernardo D, and Brunetti-Pierri N (2010) Identification of small molecules enhancing autophagic function from drug network analysis. Autophagy 6:1204–1205.
Ishii T and Mann GE (2014) Redox status in mammalian cells and stem cells during culture in vitro: critical roles of Nrf2 and cystine transporter activity in the maintenance of redox balance. Redox Biol 2:786–794.
Jang HJ, Hong EM, Kim M, Kim JH, Jang J, Park SW, Byun HW, Koh DH, Choi MH, Kae SH, et al. (2016) Simvastatin induces heme oxygenase-1 via NF-E2-related factor 2 (Nrf2) activation through ERK and PI3K/Akt pathway in colon cancer. Oncotarget 7:46219–46229.
Jansen T, Hortmann M, Oelze M, Opitz B, Steven S, Schell R, Knorr M, Karbach S, Schuhmacher S, Wenzel P, et al. (2010) Conversion of biliverdin to bilirubin by biliverdin reductase contributes to endothelial cell protection by heme oxygenase1-evidence for direct and indirect antioxidant actions of bilirubin. J Mol Cell Cardiol 49:186–195.
Jay D, Hitomi H, and Griendling KK (2006) Oxidative stress and diabetic cardiovascular complications. Free Radic Biol Med 40:183–192.
Jazwa A, Rojo AI, Innamorato NG, Hesse M, Fernández-Ruiz J, and Cuadrado A (2011) Pharmacological targeting of the transcription factor Nrf2 at the basal ganglia provides disease modifying therapy for experimental parkinsonism. Antioxid Redox Signal 14:2347–2360.
Jiang T, Huang Z, Lin Y, Zhang Z, Fang D, and Zhang DD (2010) The protective role of Nrf2 in streptozotocin-induced diabetic nephropathy. Diabetes 59:850–860.
Jiang T, Tian F, Zheng H, Whitman SA, Lin Y, Zhang Z, Zhang N, and Zhang DD (2014a) Nrf2 suppresses lupus nephritis through inhibition of oxidative injury and the NF-kB-mediated inflammatory response. Kidney Int 85:333–343.
Jiang ZY, Lu MC, Xu LL, Yang TT, Xi MY, Xu XL, Guo XK, Zhang XJ, You QD, and Sun HP (2014b) Discovery of potent Keap1-Nrf2 protein-protein interaction inhibitor based on molecular binding determinants analysis [published correction appears in J Med Chem (2014) 57:4406]. J Med Chem 57: 2736–2745.
Jiang ZY, Lu MC, and You QD (2016) Discovery and development of Kelch-like ECHassociated protein 1: Nuclear factor erythroid 2-related factor 2 (KEAP1:NRF2) protein-protein interaction inhibitors: achievements, challenges, and future directions. J Med Chem 59:10837–10858.
Jiménez-Osorio AS, González-Reyes S, García-Niño WR, Moreno-Macías H, Rodríguez-Arellano ME, Vargas-Alarcón G, Zúñiga J, Barquera R, Pedraza-Chaverri J, Meza-Espinoza JP, et al. (2017) Corrigendum to “association of nuclear factorerythroid 2-related factor 2, thioredoxin interacting protein, and heme oxygenase-1 gene polymorphisms with diabetes and obesity in Mexican patients.” Oxid Med Cell Longev 2017:7543194.
Jiménez-Osorio AS, Picazo A, González-Reyes S, Barrera-Oviedo D, Rodríguez-Arellano ME, and Pedraza-Chaverri J (2014) Nrf2 and redox status in prediabetic and diabetic patients. Int J Mol Sci 15:20290–20305.
Jnoff E, Albrecht C, Barker JJ, Barker O, Beaumont E, Bromidge S, Brookfield F, Brooks M, Bubert C, Ceska T, et al. (2014) Binding mode and structure-activity relationships around direct inhibitors of the Nrf2-Keap1 complex. ChemMedChem 9:699–705.
Jo C, Gundemir S, Pritchard S, Jin YN, Rahman I, and Johnson GV (2014) Nrf2 reduces levels of phosphorylated tau protein by inducing autophagy adaptor protein NDP52. Nat Commun 5:3496.
Johnson DA, Amirahmadi S, Ward C, Fabry Z, and Johnson JA (2010) The absence of the pro-antioxidant transcription factor Nrf2 exacerbates experimental autoimmune encephalomyelitis. Toxicol Sci 114:237–246.
Johnson DA and Johnson JA (2015) Nrf2–a therapeutic target for the treatment of neurodegenerative diseases. Free Radic Biol Med 88 (Pt B):253–267.
Johnson NM, Egner PA, Baxter VK, Sporn MB, Wible RS, Sutter TR, Groopman JD, Kensler TW, and Roebuck BD (2014) Complete protection against aflatoxin B(1)- induced liver cancer with a triterpenoid: DNA adduct dosimetry, molecular signature, and genotoxicity threshold. Cancer Prev Res (Phila) 7:658–665.
Jones RM, Desai C, Darby TM, Luo L, Wolfarth AA, Scharer CD, Ardita CS, Reedy AR, Keebaugh ES, and Neish AS (2015) Lactobacilli modulate epithelial cytoprotection through the Nrf2 pathway. Cell Reports 12:1217–1225.
Jung KA, Lee S, and Kwak MK (2017) NFE2L2/NRF2 activity is linked to mitochondria and AMP-activated protein kinase signaling in cancers through miR181c/mitochondria-encoded cytochrome c oxidase regulation. Antioxid Redox Signal 27:945–961.
Kaidery NA, Banerjee R, Yang L, Smirnova NA, Hushpulian DM, Liby KT, Williams CR, Yamamoto M, Kensler TW, Ratan RR, et al. (2013) Targeting Nrf2-mediated gene transcription by extremely potent synthetic triterpenoids attenuates dopaminergic neurotoxicity in the MPTP mouse model of Parkinson’s disease. Antioxid Redox Signal 18:139–157.
Kaisar MA, Villalba H, Prasad S, Liles T, Sifat AE, Sajja RK, Abbruscato TJ, and Cucullo L (2017) Offsetting the impact of smoking and e-cigarette vaping on the cerebrovascular system and stroke injury: is Metformin a viable countermeasure? Redox Biol 13:353–362.
Kastrati I, Siklos MI, Calderon-Gierszal EL, El-Shennawy L, Georgieva G, Thayer EN, Thatcher GR, and Frasor J (2016) Dimethyl fumarate inhibits the nuclear factor kB pathway in breast cancer cells by covalent modification of p65 protein. J Biol Chem 291:3639–3647.
Kelleni MT, Amin EF, and Abdelrahman AM (2015) Effect of metformin and sitagliptin on doxorubicin-induced cardiotoxicity in rats: impact of oxidative stress, inflammation, and apoptosis. J Toxicol 2015:424813. Kensler TW, Egner PA, Agyeman AS, Visvanathan K, Groopman JD, Chen JG, Chen TY, Fahey JW, and Talalay P (2013) Keap1-nrf2 signaling: a target for cancer prevention by sulforaphane. Top Curr Chem 329:163–177.
Khan I, Tantray MA, Alam MS, and Hamid H (2017) Natural and synthetic bioactive inhibitors of glycogen synthase kinase. Eur J Med Chem 125:464–477. Kim JE, You DJ, Lee C, Ahn C, Seong JY, and Hwang JI (2010a) Suppression of NF-kappaB signaling by KEAP1 regulation of IKKbeta activity through autophagic degradation and inhibition of phosphorylation. Cell Signal 22:1645–1654.
Kim T, Kim YJ, Han IH, Lee D, Ham J, Kang KS, and Lee JW (2015) The synthesis of sulforaphane analogues and their protection effect against cisplatin induced cytotoxicity in kidney cells. Bioorg Med Chem Lett 25:62–66.
Kim YR, Oh JE, Kim MS, Kang MR, Park SW, Han JY, Eom HS, Yoo NJ, and Lee SH (2010b) Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin. J Pathol 220:446–451.
Kitagishi Y and Matsuda S (2013) Redox regulation of tumor suppressor PTEN in cancer and aging (Review). Int J Mol Med 31:511–515.
Kitsak M, Sharma A, Menche J, Guney E, Ghiassian SD, Loscalzo J, and Barabási AL (2016) Tissue specificity of human disease module. Sci Rep 6:35241. Kobayashi EH, Suzuki T, Funayama R, Nagashima T, Hayashi M, Sekine H, Tanaka N, Moriguchi T, Motohashi H, Nakayama K, et al. (2016) Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat Commun 7:11624.
Kocer D, Bayram F, and Diri H (2014) The effects of metformin on endothelial dysfunction, lipid metabolism and oxidative stress in women with polycystic ovary syndrome. Gynecol Endocrinol 30:367–371.
Komatsu M, Kurokawa H, Waguri S, Taguchi K, Kobayashi A, Ichimura Y, Sou YS, Ueno I, Sakamoto A, Tong KI, et al. (2010) The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat Cell Biol 12:213–223.
Kong X, Thimmulappa R, Craciun F, Harvey C, Singh A, Kombairaju P, Reddy SP, Remick D, and Biswal S (2011) Enhancing Nrf2 pathway by disruption of Keap1 in myeloid leukocytes protects against sepsis. Am J Respir Crit Care Med 184: 928–938.
Kruger AL, Peterson SJ, Schwartzman ML, Fusco H, McClung JA, Weiss M, Shenouda S, Goodman AI, Goligorsky MS, Kappas A, et al. (2006) Up-regulation of heme oxygenase provides vascular protection in an animal model of diabetes through its antioxidant and antiapoptotic effects. J Pharmacol Exp Ther 319: 1144–1152.
Kruse ML, Friedrich M, Arlt A, Röcken C, Egberts JH, Sebens S, and Schäfer H (2016) Colonic lamina propria inflammatory cells from patients with IBD induce the nuclear factor-E2 related factor-2 thereby leading to greater proteasome activity and apoptosis protection in human colonocytes. Inflamm Bowel Dis 22: 2593–2606.
Kuang X, Scofield VL, Yan M, Stoica G, Liu N, and Wong PK (2009) Attenuation of oxidative stress, inflammation and apoptosis by minocycline prevents retrovirusinduced neurodegeneration in mice. Brain Res 1286:174–184.
Kurinna S and Werner S (2015) NRF2 and microRNAs: new but awaited relations. Biochem Soc Trans 43:595–601.
Kwak JY, Takeshige K, Cheung BS, and Minakami S (1991) Bilirubin inhibits the activation of superoxide-producing NADPH oxidase in a neutrophil cell-free system. Biochim Biophys Acta 1076:369–373.
Lamb J, Crawford ED, Peck D, Modell JW, Blat IC, Wrobel MJ, Lerner J, Brunet JP, Subramanian A, Ross KN, et al. (2006) The connectivity map: using geneexpression signatures to connect small molecules, genes, and disease. Science 313: 1929–1935.
Lastres-Becker I, García-Yagüe AJ, Scannevin RH, Casarejos MJ, Kügler S, Rábano A, and Cuadrado A (2016) Repurposing the NRF2 activator dimethyl fumarate as therapy against synucleinopathy in Parkinson’s disease. Antioxid Redox Signal 25: 61–77.
Lastres-Becker I, Innamorato NG, Jaworski T, Rábano A, Kügler S, Van Leuven F, and Cuadrado A (2014) Fractalkine activates NRF2/NFE2L2 and heme oxygenase 1 to restrain tauopathy-induced microgliosis. Brain 137:78–91.
Lastres-Becker I, Ulusoy A, Innamorato NG, Sahin G, Rábano A, Kirik D, and Cuadrado A (2012) a-Synuclein expression and Nrf2 deficiency cooperate to aggravate protein aggregation, neuronal death and inflammation in early-stage Parkinson’s disease. Hum Mol Genet 21:3173–3192.
Lee D, Xu IM, Chiu DK, Lai RK, Tse AP, Lan Li L, Law CT, Tsang FH, Wei LL, Chan CY, et al. (2017) Folate cycle enzyme MTHFD1L confers metabolic advantages in hepatocellular carcinoma. J Clin Invest 127:1856–1872.
Lee DF, Kuo HP, Liu M, Chou CK, Xia W, Du Y, Shen J, Chen CT, Huo L, Hsu MC, et al. (2009) KEAP1 E3 ligase-mediated downregulation of NF-kappaB signaling by targeting IKKbeta. Mol Cell 36:131–140.
Lee SR, Yang KS, Kwon J, Lee C, Jeong W, and Rhee SG (2002) Reversible inactivation of the tumor suppressor PTEN by H2O2. J Biol Chem 277:20336–20342.
Li B, Cui W, Liu J, Li R, Liu Q, Xie XH, Ge XL, Zhang J, Song XJ, Wang Y, et al. (2013) Sulforaphane ameliorates the development of experimental autoimmune encephalomyelitis by antagonizing oxidative stress and Th17-related inflammation in mice. Exp Neurol 250:239–249.
Li J, Stein TD, and Johnson JA (2004) Genetic dissection of systemic autoimmune disease in Nrf2-deficient mice. Physiol Genomics 18:261–272.
Licht-Mayer S, Wimmer I, Traffehn S, Metz I, Brück W, Bauer J, Bradl M, and Lassmann H (2015) Cell type-specific Nrf2 expression in multiple sclerosis lesions. Acta Neuropathol 130:263–277.
Limonciel A and Jennings P (2014) A review of the evidence that ochratoxin A is an Nrf2 inhibitor: implications for nephrotoxicity and renal carcinogenicity. Toxins (Basel) 6:371–379.
Lin CH, Lin HI, Chen ML, Lai TT, Cao LP, Farrer MJ, Wu RM, and Chien CT (2016) Lovastatin protects neurite degeneration in LRRK2-G2019S parkinsonism through activating the Akt/Nrf pathway and inhibiting GSK3b activity. Hum Mol Genet 25: 1965–1978.
Lin SX, Lisi L, Dello Russo C, Polak PE, Sharp A, Weinberg G, Kalinin S, and Feinstein DL (2011) The anti-inflammatory effects of dimethyl fumarate in astrocytes involve glutathione and haem oxygenase-1. ASN Neuro 3:75–84.
Lin W, Wu RT, Wu T, Khor TO, Wang H, and Kong AN (2008) Sulforaphane suppressed LPS-induced inflammation in mouse peritoneal macrophages through Nrf2 dependent pathway. Biochem Pharmacol 76:967–973.
Linker RA, Lee DH, Ryan S, van Dam AM, Conrad R, Bista P, Zeng W, Hronowsky X, Buko A, Chollate S, et al. (2011) Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the Nrf2 antioxidant pathway. Brain 134: 678–692.
Liu GH, Qu J, and Shen X (2008) NF-kappaB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim Biophys Acta 1783:713–727.
Liu TS, Pei YH, Peng YP, Chen J, Jiang SS, and Gong JB (2014) Oscillating high glucose enhances oxidative stress and apoptosis in human coronary artery endothelial cells. J Endocrinol Invest 37:645–651.
Lo SC, Li X, Henzl MT, Beamer LJ, and Hannink M (2006) Structure of the Keap1: Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J 25: 3605–3617.
LoGerfo A, Chico L, Borgia L, Petrozzi L, Rocchi A, D’Amelio A, Carlesi C, Caldarazzo Ienco E, Mancuso M, and Siciliano G (2014) Lack of association between nuclear factor erythroid-derived 2-like 2 promoter gene polymorphisms and oxidative stress biomarkers in amyotrophic lateral sclerosis patients. Oxid Med Cell Longev 2014:432626.
Lu K, Alcivar AL, Ma J, Foo TK, Zywea S, Mahdi A, Huo Y, Kensler TW, Gatza ML, and Xia B (2017) NRF2 induction supporting breast cancer cell survival is enabled by oxidative stress-induced DPP3-KEAP1 interaction. Cancer Res 77: 2881–2892.
Lu MC, Jiao Q, Liu T, Tan SJ, Zhou HS, You QD, and Jiang ZY (2018) Discovery of a head-to-tail cyclic peptide as the Keap1-Nrf2 protein-protein interaction inhibitor with high cell potency. Eur J Med Chem 143:1578–1589.
Lv H, Liu Q, Wen Z, Feng H, Deng X, and Ci X (2017) Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute lung injury via induction of AMPK/GSK3bNrf2 signal axis. Redox Biol 12:311–324.
Ma Q (2013) Role of nrf2 in oxidative stress and toxicity. Annu Rev Pharmacol Toxicol 53:401–426.
Ma Q, Battelli L, and Hubbs AF (2006) Multiorgan autoimmune inflammation, enhanced lymphoproliferation, and impaired homeostasis of reactive oxygen species in mice lacking the antioxidant-activated transcription factor Nrf2. Am J Pathol 168:1960–1974.
Maicas N, Ferrándiz ML, Brines R, Ibáñez L, Cuadrado A, Koenders MI, van den Berg WB, and Alcaraz MJ (2011) Deficiency of Nrf2 accelerates the effector phase of arthritis and aggravates joint disease. Antioxid Redox Signal 15:889–901.
Manna A, De Sarkar S, De S, Bauri AK, Chattopadhyay S, and Chatterjee M (2015) The variable chemotherapeutic response of malabaricone-A in leukemic and solid tumor cell lines depends on the degree of redox imbalance. Phytomedicine 22: 713–723.
Maqbool M and Hoda N (2017) GSK3 inhibitors in the therapeutic development of diabetes, cancer and neurodegeneration: past, present and future. Curr Pharm Des 23:4332–4350.
Marcotte D, Zeng W, Hus JC, McKenzie A, Hession C, Jin P, Bergeron C, Lugovskoy A, Enyedy I, Cuervo H, et al. (2013) Small molecules inhibit the interaction of Nrf2 and the Keap1 Kelch domain through a non-covalent mechanism. Bioorg Med Chem 21:4011–4019.
Markart P, Luboeinski T, Korfei M, Schmidt R, Wygrecka M, Mahavadi P, Mayer K, Wilhelm J, Seeger W, Guenther A, et al. (2009) Alveolar oxidative stress is associated with elevated levels of nonenzymatic low-molecular-weight antioxidants in patients with different forms of chronic fibrosing interstitial lung diseases. Antioxid Redox Signal 11:227–240.
Markowicz-Piasecka M, Sikora J, Szyd?owska A, Skupien A, Mikiciuk-Olasik E, and Huttunen KM (2017) Metformin - a future therapy for neurodegenerative diseases: theme: drug discovery, development and delivery in Alzheimer’s disease: guest editor: Davide Brambilla. Pharm Res 34:2614–2627.
Marrone G, Maeso-Díaz R, García-Cardena G, Abraldes JG, García-Pagán JC, Bosch J, and Gracia-Sancho J (2015) KLF2 exerts antifibrotic and vasoprotective effects in cirrhotic rat livers: behind the molecular mechanisms of statins. Gut 64: 1434–1443.
Martín-de-Saavedra MD, Budni J, Cunha MP, Gómez-Rangel V, Lorrio S, Del Barrio L, Lastres-Becker I, Parada E, Tordera RM, Rodrigues AL, et al. (2013) Nrf2 participates in depressive disorders through an anti-inflammatory mechanism. Psychoneuroendocrinology 38:2010–2022.
Marzec JM, Christie JD, Reddy SP, Jedlicka AE, Vuong H, Lanken PN, Aplenc R, Yamamoto T, Yamamoto M, Cho HY, et al. (2007) Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J 21:2237–2246.
McCubrey JA, Steelman LS, Bertrand FE, Davis NM, Sokolosky M, Abrams SL, Montalto G, D’Assoro AB, Libra M, Nicoletti F, et al. (2014) GSK-3 as potential target for therapeutic intervention in cancer. Oncotarget 5:2881–2911.
McGuire VA, Ruiz-Zorrilla Diez T, Emmerich CH, Strickson S, Ritorto MS, Sutavani RV, Weib A, Houslay KF, Knebel A, Meakin PJ, et al. (2016) Dimethyl fumarate blocks pro-inflammatory cytokine production via inhibition of TLR induced M1 and K63 ubiquitin chain formation. Sci Rep 6:31159.
Meakin PJ, Chowdhry S, Sharma RS, Ashford FB, Walsh SV, McCrimmon RJ, Dinkova-Kostova AT, Dillon JF, Hayes JD, and Ashford ML (2014) Susceptibility of Nrf2-null mice to steatohepatitis and cirrhosis upon consumption of a high-fat diet is associated with oxidative stress, perturbation of the unfolded protein response, and disturbance in the expression of metabolic enzymes but not with insulin resistance. Mol Cell Biol 34:3305–3320.
Meissner M, Doll M, Hrgovic I, Reichenbach G, König V, Hailemariam-Jahn T, Gille J, and Kaufmann R (2011) Suppression of VEGFR2 expression in human endothelial cells by dimethylfumarate treatment: evidence for anti-angiogenic action. J Invest Dermatol 131:1356–1364.
Menche J, Sharma A, Kitsak M, Ghiassian SD, Vidal M, Loscalzo J, and Barabási AL (2015) Disease networks: uncovering disease-disease relationships through the incomplete interactome. Science 347:1257601.
Milkovic L, Zarkovic N, and Saso L (2017) Controversy about pharmacological modulation of Nrf2 for cancer therapy. Redox Biol 12:727–732.
Mitsuishi Y, Taguchi K, Kawatani Y, Shibata T, Nukiwa T, Aburatani H, Yamamoto M, and Motohashi H (2012) Nrf2 redirects glucose and glutamine into anabolic pathways in metabolic reprogramming. Cancer Cell 22:66–79.
Moon SJ, Park JS, Woo YJ, Lim MA, Kim SM, Lee SY, Kim EK, Lee HJ, Lee WS, Park SH, et al. (2014) Rebamipide suppresses collagen-induced arthritis through reciprocal regulation of th17/treg cell differentiation and heme oxygenase 1 induction. Arthritis Rheumatol 66:874–885.
Morgan MJ and Liu ZG (2011) Crosstalk of reactive oxygen species and NF-kB signaling. Cell Res 21:103–115.
Morgan PE, Sturgess AD, Hennessy A, and Davies MJ (2007) Serum protein oxidation and apolipoprotein CIII levels in people with systemic lupus erythematosus with and without nephritis. Free Radic Res 41:1301–1312.
Mrowietz U, Christophers E, and Altmeyer P (1998) Treatment of psoriasis with fumaric acid esters: results of a prospective multicentre study: German Multicentre study. Br J Dermatol 138:456–460.
Mrowietz U, Szepietowski JC, Loewe R, van de Kerkhof P, Lamarca R, Ocker WG, Tebbs VM, and Pau-Charles I (2017) Efficacy and safety of LAS41008 (dimethyl fumarate) in adults with moderate-to-severe chronic plaque psoriasis: a randomized, double-blind, Fumaderm®- and placebo-controlled trial (BRIDGE). Br J Dermatol 176:615–623.
Na LX, Li Y, Pan HZ, Zhou XL, Sun DJ, Meng M, Li XX, and Sun CH (2013) Curcuminoids exert glucose-lowering effect in type 2 diabetes by decreasing serum free fatty acids: a double-blind, placebo-controlled trial. Mol Nutr Food Res 57: 1569–1577.
Napetschnig J and Wu H (2013) Molecular basis of NF-kB signaling. Annu Rev Biophys 42:443–468.
Nasiri HR, Linge S, and Ullmann D (2016) Thermodynamic profiling of inhibitors of Nrf2:Keap1 interactions. Bioorg Med Chem Lett 26:526–529.
Natarajan VT, Singh A, Kumar AA, Sharma P, Kar HK, Marrot L, Meunier JR, Natarajan K, Rani R, and Gokhale RS (2010) Transcriptional upregulation of Nrf2- dependent phase II detoxification genes in the involved epidermis of vitiligo vulgaris. J Invest Dermatol 130:2781–2789.
Nef HM, Möllmann H, Troidl C, Kostin S, Böttger T, Voss S, Hilpert P, Krause N, Weber M, Rolf A, et al. (2008) Expression profiling of cardiac genes in Tako-Tsubo cardiomyopathy: insight into a new cardiac entity. J Mol Cell Cardiol 44:395–404.
Nesti L and Natali A (2017) Metformin effects on the heart and the cardiovascular system: a review of experimental and clinical data. Nutr Metab Cardiovasc Dis 27: 657–669.
Ockenfels HM, Schultewolter T, Ockenfels G, Funk R, and Goos M (1998) The antipsoriatic agent dimethylfumarate immunomodulates T-cell cytokine secretion and inhibits cytokines of the psoriatic cytokine network. Br J Dermatol 139: 390–395.
Okano Y, Nezu U, Enokida Y, Lee MT, Kinoshita H, Lezhava A, Hayashizaki Y, Morita S, Taguri M, Ichikawa Y, et al. (2013) SNP (-617C.A) in ARE-like loci of the NRF2 gene: a new biomarker for prognosis of lung adenocarcinoma in Japanese non-smoking women. PLoS One 8:e73794.
Olayanju A, Copple IM, Bryan HK, Edge GT, Sison RL, Wong MW, Lai ZQ, Lin ZX, Dunn K, Sanderson CM, et al. (2015) Brusatol provokes a rapid and transient inhibition of Nrf2 signaling and sensitizes mammalian cells to chemical toxicity: implications for therapeutic targeting of Nrf2. Free Radic Biol Med 78:202–212.
Onderdijk AJ, Balak DM, Baerveldt EM, Florencia EF, Kant M, Laman JD, van IJcken WF, Racz E, de Ridder D, Thio HB, et al. (2014) Regulated genes in psoriatic skin during treatment with fumaric acid esters. Br J Dermatol 171:732–741.
Onyiah JC, Sheikh SZ, Maharshak N, Otterbein LE, and Plevy SE (2014) Heme oxygenase-1 and carbon monoxide regulate intestinal homeostasis and mucosal immune responses to the enteric microbiota. Gut Microbes 5:220–224.
Orena S, Owen J, Jin F, Fabian M, Gillitt ND, and Zeisel SH (2015) Extracts of fruits and vegetables activate the antioxidant response element in IMR-32 cells. J Nutr 145:2006–2011.
Owen MR, Doran E, and Halestrap AP (2000) Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem J 348:607–614.
Padmanabhan B, Tong KI, Ohta T, Nakamura Y, Scharlock M, Ohtsuji M, Kang MI, Kobayashi A, Yokoyama S, and Yamamoto M (2006) Structural basis for defects of Keap1 activity provoked by its point mutations in lung cancer. Mol Cell 21: 689–700.
Pajares M, Cuadrado A, and Rojo AI (2017) Modulation of proteostasis by transcription factor NRF2 and impact in neurodegenerative diseases. Redox Biol 11: 543–553.
Pajares M, Jiménez-Moreno N, Dias IH, Debelec B, Vucetic M, Fladmark KE, Basaga H, Ribaric S, Milisav I, and Cuadrado A (2015) Redox control of protein degradation. Redox Biol 6:409–420.
Pajares M, Jiménez-Moreno N, García-Yagüe AJ, Escoll M, de Ceballos ML, Van Leuven F, Rábano A, Yamamoto M, Rojo AI, and Cuadrado A (2016) Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 12: 1902–1916.
Palomo V and Martinez A (2017) Glycogen synthase kinase 3 (GSK-3) inhibitors: a patent update (2014-2015). Expert Opin Ther Pat 27:657–666.
Panieri E and Santoro MM (2016) ROS homeostasis and metabolism: a dangerous liason in cancer cells. Cell Death Dis 7:e2253.
Pantan R, Tocharus J, Suksamrarn A, and Tocharus C (2016) Synergistic effect of atorvastatin and cyanidin-3-glucoside on angiotensin II-induced inflammation in vascular smooth muscle cells. Exp Cell Res 342:104–112.
Pareek TK, Belkadi A, Kesavapany S, Zaremba A, Loh SL, Bai L, Cohen ML, Meyer C, Liby KT, Miller RH, et al. (2011) Triterpenoid modulation of IL-17 and Nrf-2 expression ameliorates neuroinflammation and promotes remyelination in autoimmune encephalomyelitis. Sci Rep 1:201.
Park SY, Lee SW, Shin HK, Chung WT, Lee WS, Rhim BY, Hong KW, and Kim CD (2010) Cilostazol enhances apoptosis of synovial cells from rheumatoid arthritis patients with inhibition of cytokine formation via Nrf2-linked heme oxygenase 1 induction. Arthritis Rheum 62:732–741.
Pedruzzi LM, Cardozo LF, Daleprane JB, Stockler-Pinto MB, Monteiro EB, Leite M Jr, Vaziri ND, and Mafra D (2015) Systemic inflammation and oxidative stress in hemodialysis patients are associated with down-regulation of Nrf2. J Nephrol 28: 495–501.
Peng H, Guerau-de-Arellano M, Mehta VB, Yang Y, Huss DJ, Papenfuss TL, LovettRacke AE, and Racke MK (2012) Dimethyl fumarate inhibits dendritic cell maturation via nuclear factor kB (NF-kB) and extracellular signal-regulated kinase 1 and 2 (ERK1/2) and mitogen stress-activated kinase 1 (MSK1) signaling. J Biol Chem 287:28017–28026.
Pergola PE, Raskin P, Toto RD, Meyer CJ, Huff JW, Grossman EB, Krauth M, Ruiz S, Audhya P, Christ-Schmidt H, et al.; BEAM Study Investigators (2011) Bardoxolone methyl and kidney function in CKD with type 2 diabetes. N Engl J Med 365:327–336.
Pilar Valdecantos M, Prieto-Hontoria PL, Pardo V, Módol T, Santamaría B, Weber M, Herrero L, Serra D, Muntané J, Cuadrado A, et al. (2015) Essential role of Nrf2 in the protective effect of lipoic acid against lipoapoptosis in hepatocytes. Free Radic Biol Med 84:263–278.
Piñero J, Bravo À, Queralt-Rosinach N, Gutiérrez-Sacristán A, Deu-Pons J, Centeno E, García-García J, Sanz F, and Furlong LI (2017) DisGeNET: a comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res 45:D833–D839.
Pitha-Rowe I, Liby K, Royce D, and Sporn M (2009) Synthetic triterpenoids attenuate cytotoxic retinal injury: cross-talk between Nrf2 and PI3K/AKT signaling through inhibition of the lipid phosphatase PTEN. Invest Ophthalmol Vis Sci 50: 5339–5347.
Qiu P, Man S, Li J, Liu J, Zhang L, Yu P, and Gao W (2016) Overdose intake of curcumin initiates the unbalanced state of bodies. J Agric Food Chem 64: 2765–2771.
Rachakonda G, Xiong Y, Sekhar KR, Stamer SL, Liebler DC, and Freeman ML (2008) Covalent modification at Cys151 dissociates the electrophile sensor Keap1 from the ubiquitin ligase CUL3. Chem Res Toxicol 21:705–710.
Rada P, Rojo AI, Chowdhry S, McMahon M, Hayes JD, and Cuadrado A (2011) SCF/ beta-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner. Mol Cell Biol 31:1121–1133.
Rada P, Rojo AI, Evrard-Todeschi N, Innamorato NG, Cotte A, Jaworski T, TobónVelasco JC, Devijver H, García-Mayoral MF, Van Leuven F, et al. (2012) Structural and functional characterization of Nrf2 degradation by the glycogen synthase kinase 3/b-TrCP axis. Mol Cell Biol 32:3486–3499.
Ramadori P, Drescher H, Erschfeld S, Schumacher F, Berger C, Fragoulis A, Schenkel J, Kensler TW, Wruck CJ, Trautwein C, et al. (2016) Hepatocyte-specific Keap1 deletion reduces liver steatosis but not inflammation during non-alcoholic steatohepatitis development. Free Radic Biol Med 91:114–126.
Ramsey CP, Glass CA, Montgomery MB, Lindl KA, Ritson GP, Chia LA, Hamilton RL, Chu CT, and Jordan-Sciutto KL (2007) Expression of Nrf2 in neurodegenerative diseases. J Neuropathol Exp Neurol 66:75–85.
Rangasamy T, Cho CY, Thimmulappa RK, Zhen L, Srisuma SS, Kensler TW, Yamamoto M, Petrache I, Tuder RM, and Biswal S (2004) Genetic ablation of Nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J Clin Invest 114:1248–1259.
Ranjan N, Fulcrand G, King A, Brown J, Jiang X, Leng F, and Arya DP (2014) Selective inhibition of bacterial topoisomerase I by alkynyl-bisbenzimidazoles. MedChemComm 5:816–825.
Reccia I, Kumar J, Akladios C, Virdis F, Pai M, Habib N, and Spalding D (2017) Nonalcoholic fatty liver disease: a sign of systemic disease. Metabolism 72:94–108.
Ren D, Villeneuve NF, Jiang T, Wu T, Lau A, Toppin HA, and Zhang DD (2011) Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proc Natl Acad Sci USA 108:1433–1438.
Rena G, Hardie DG, and Pearson ER (2017) The mechanisms of action of metformin. Diabetologia 60:1577–1585.
Reuter S, Gupta SC, Chaturvedi MM, and Aggarwal BB (2010) Oxidative stress, inflammation, and cancer: how are they linked? Free Radic Biol Med 49:1603–1616.
Richardson BG, Jain AD, Speltz TE, and Moore TW (2015) Non-electrophilic modulators of the canonical Keap1/Nrf2 pathway. Bioorg Med Chem Lett 25:2261–2268.
Roberts CA, Dickinson AK, and Taams LS (2015) The interplay between monocytes/ macrophages and CD4(+) T cell subsets in rheumatoid arthritis. Front Immunol 6: 571.
Rojo AI, Innamorato NG, Martín-Moreno AM, De Ceballos ML, Yamamoto M, and Cuadrado A (2010) Nrf2 regulates microglial dynamics and neuroinflammation in experimental Parkinson’s disease. Glia 58:588–598.
Rojo AI, McBean G, Cindric M, Egea J, López MG, Rada P, Zarkovic N, and Cuadrado A (2014a) Redox control of microglial function: molecular mechanisms and functional significance. Antioxid Redox Signal 21:1766–1801.
Rojo AI, Rada P, Mendiola M, Ortega-Molina A, Wojdyla K, Rogowska-Wrzesinska A, Hardisson D, Serrano M, and Cuadrado A (2014b) The PTEN/NRF2 axis promotes human carcinogenesis. Antioxid Redox Signal 21:2498–2514.
Rushworth SA, Zaitseva L, Murray MY, Shah NM, Bowles KM, and MacEwan DJ (2012) The high Nrf2 expression in human acute myeloid leukemia is driven by NF-kB and underlies its chemo-resistance. Blood 120:5188–5198.
Ryoo IG, Lee SH, and Kwak MK (2016) Redox modulating NRF2: a potential mediator of cancer stem cell resistance. Oxid Med Cell Longev 2016:2428153.
Saddawi-Konefka R, Seelige R, Gross ET, Levy E, Searles SC, Washington A Jr, Santosa EK, Liu B, O’Sullivan TE, Harismendy O, et al. (2016) Nrf2 induces IL-17D to mediate tumor and virus surveillance. Cell Reports 16:2348–2358.
Saito R, Suzuki T, Hiramoto K, Asami S, Naganuma E, Suda H, Iso T, Yamamoto H, Morita M, Baird L, et al. (2015) Characterizations of three major cysteine sensors of Keap1 in stress response. Mol Cell Biol 36:271–284.
Saracino AM and Orteu CH (2017) Severe recalcitrant cutaneous manifestations in systemic lupus erythematosus successfully treated with fumaric acid esters. Br J Dermatol 176:472–480.
Sato M, Aoki T, Inoue H, Tanaka T, and Kunishima N (2013) Keap1 Protein Binding Compound, Cristal of Complex Between the Same and Keap1 Protein, and Method for Producing the Same, Toray Industries, Tokyo. Sato N, Takasaka N, Yoshida M, Tsubouchi K, Minagawa S, Araya J, Saito N, Fujita Y, Kurita Y, Kobayashi K, et al. (2016) Metformin attenuates lung fibrosis development via NOX4 suppression. Respir Res 17:107.
Satoh T, McKercher SR, and Lipton SA (2013) Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic Biol Med 65:645–657.
Scannevin RH, Chollate S, Jung MY, Shackett M, Patel H, Bista P, Zeng W, Ryan S, Yamamoto M, Lukashev M, et al. (2012) Fumarates promote cytoprotection of central nervous system cells against oxidative stress via the nuclear factor (erythroid-derived 2)-like 2 pathway. J Pharmacol Exp Ther 341:274–284.
Schilling S, Goelz S, Linker R, Luehder F, and Gold R (2006) Fumaric acid esters are effective in chronic experimental autoimmune encephalomyelitis and suppress macrophage infiltration. Clin Exp Immunol 145:101–107.
Schimrigk S, Brune N, Hellwig K, Lukas C, Bellenberg B, Rieks M, Hoffmann V, Pohlau D, and Przuntek H (2006) Oral fumaric acid esters for the treatment of active multiple sclerosis: an open-label, baseline-controlled pilot study. Eur J Neurol 13:604–610.
Schipper HM, Song W, Zukor H, Hascalovici JR, and Zeligman D (2009) Heme oxygenase-1 and neurodegeneration: expanding frontiers of engagement. J Neurochem 110:469–485.
Schuiveling M, Vazirpanah N, Radstake TRDJ, Zimmermann M, and Broen JCA (2017) Metformin, a new era for an old drug in the treatment of immune mediated disease? Curr Drug Targets [published ahead of print]. Schulze-Topphoff U, Varrin-Doyer M, Pekarek K, Spencer CM, Shetty A, Sagan SA, Cree BA, Sobel RA, Wipke BT, Steinman L, et al. (2016) Dimethyl fumarate treatment induces adaptive and innate immune modulation independent of Nrf2. Proc Natl Acad Sci USA 113:4777–4782.
Schumacker PT (2006) Reactive oxygen species in cancer cells: live by the sword, die by the sword. Cancer Cell 10:175–176.
Sha LK, Sha W, Kuchler L, Daiber A, Giegerich AK, Weigert A, Knape T, Snodgrass R, Schröder K, Brandes RP, et al. (2015) Loss of Nrf2 in bone marrow-derived macrophages impairs antigen-driven CD8(+) T cell function by limiting GSH and Cys availability. Free Radic Biol Med 83:77–88.
Shanmugam G, Narasimhan M, Conley RL, Sairam T, Kumar A, Mason RP, Sankaran R, Hoidal JR, and Rajasekaran NS (2017a) Chronic endurance exercise impairs cardiac structure and function in middle-aged mice with impaired Nrf2 signaling. Front Physiol 8:268.
Shanmugam G, Narasimhan M, Tamowski S, Darley-Usmar V, and Rajasekaran NS (2017b) Constitutive activation of Nrf2 induces a stable reductive state in the mouse myocardium. Redox Biol 12:937–945.
Sharma A, Rizky L, Stefanovic N, Tate M, Ritchie RH, Ward KW, and de Haan JB (2017) The nuclear factor (erythroid-derived 2)-like 2 (Nrf2) activator dh404 protects against diabetes-induced endothelial dysfunction. Cardiovasc Diabetol 16:33.
Shibata T, Kokubu A, Saito S, Narisawa-Saito M, Sasaki H, Aoyagi K, Yoshimatsu Y, Tachimori Y, Kushima R, Kiyono T, et al. (2011) NRF2 mutation confers malignant potential and resistance to chemoradiation therapy in advanced esophageal squamous cancer. Neoplasia 13:864–873.
Shimizu T, Inoue K, Hachiya H, Shibuya N, Aoki T, and Kubota K (2016) Accumulation of phosphorylated p62 is associated with NF-E2-related factor 2 activation in hepatocellular carcinoma. J Hepatobiliary Pancreat Sci 23:467–471.
Shimoyama Y, Mitsuda Y, Tsuruta Y, Hamajima N, and Niwa T (2014) Polymorphism of Nrf2, an antioxidative gene, is associated with blood pressure and cardiovascular mortality in hemodialysis patients. Int J Med Sci 11:726–731.
Shimozono R, Asaoka Y, Yoshizawa Y, Aoki T, Noda H, Yamada M, Kaino M, and Mochizuki H (2013) Nrf2 activators attenuate the progression of nonalcoholic steatohepatitis-related fibrosis in a dietary rat model. Mol Pharmacol 84:62–70.
Signorelli SS, Li Volsi G, Fiore V, Mangiafico M, Barbagallo I, Parenti R, Rizzo M, and Li Volti G (2016) Plasma heme oxygenase-1 is decreased in peripheral artery disease patients. Mol Med Rep 14:3459–3463.
Silva T, Reis J, Teixeira J, and Borges F (2014) Alzheimer’s disease, enzyme targets and drug discovery struggles: from natural products to drug prototypes. Ageing Res Rev 15:116–145.
Singh A, Misra V, Thimmulappa RK, Lee H, Ames S, Hoque MO, Herman JG, Baylin SB, Sidransky D, Gabrielson E, et al. (2006) Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med 3:e420.
Singh A, Venkannagari S, Oh KH, Zhang YQ, Rohde JM, Liu L, Nimmagadda S, Sudini K, Brimacombe KR, Gajghate S, et al. (2016) Small molecule inhibitor of NRF2 selectively intervenes therapeutic resistance in KEAP1-deficient NSCLC tumors. ACS Chem Biol 11:3214–3225.
Siomek A (2012) NF-kB signaling pathway and free radical impact. Acta Biochim Pol 59:323–331.
Song P, Li K, Liu L, Wang X, Jian Z, Zhang W, Wang G, Li C, and Gao T (2016) Genetic polymorphism of the Nrf2 promoter region is associated with vitiligo risk in Han Chinese populations. J Cell Mol Med 20:1840–1850.
Stelzer G, Rosen N, Plaschkes I, Zimmerman S, Twik M, Fishilevich S, Stein TI, Nudel R, Lieder I, Mazor Y, et al. (2016) The GeneCards suite: from gene data mining to disease genome sequence analyses. Curr Protoc Bioinformatics 54: 1.30.1–1.30.33.
Stocker R, Yamamoto Y, McDonagh AF, Glazer AN, and Ames BN (1987) Bilirubin is an antioxidant of possible physiological importance. Science 235:1043–1046.
Strom J and Chen QM (2017) Loss of Nrf2 promotes rapid progression to heart failure following myocardial infarction. Toxicol Appl Pharmacol 327:52–58.
Sun H, Zhu J, Lin H, Gu K, and Feng F (2017) Recent progress in the development of small molecule Nrf2 modulators: a patent review (2012-2016). Expert Opin Ther Pat 27:763–785.
Taguchi K and Yamamoto M (2017) The KEAP1-NRF2 system in cancer. Front Oncol 7:85.
Tahvili S, Zandieh B, and Amirghofran Z (2015) The effect of dimethyl fumarate on gene expression and the level of cytokines related to different T helper cell subsets in peripheral blood mononuclear cells of patients with psoriasis. Int J Dermatol 54: e254–e260.
Takahashi Y, Kobayashi Y, Kawata K, Kawamura K, Sumiyoshi S, Noritake H, Watanabe S, Chida T, Souda K, Sakaguchi T, et al. (2014) Does hepatic oxidative stress enhance activation of nuclear factor-E2-related factor in patients with nonalcoholic steatohepatitis? Antioxid Redox Signal 20:538–543.
Tan SM, Sharma A, Stefanovic N, Yuen DY, Karagiannis TC, Meyer C, Ward KW, Cooper ME, and de Haan JB (2014) Derivative of bardoxolone methyl, dh404, in an inverse dose-dependent manner lessens diabetes-associated atherosclerosis and improves diabetic kidney disease. Diabetes 63:3091–3103.
Tang X, Lin CC, Spasojevic I, Iversen ES, Chi JT, and Marks JR (2014) A joint analysis of metabolomics and genetics of breast cancer. Breast Cancer Res 16:415.
Tarumoto T, Nagai T, Ohmine K, Miyoshi T, Nakamura M, Kondo T, Mitsugi K, Nakano S, Muroi K, Komatsu N, et al. (2004) Ascorbic acid restores sensitivity to imatinib via suppression of Nrf2-dependent gene expression in the imatinibresistant cell line. Exp Hematol 32:375–381.
Thimmulappa RK, Lee H, Rangasamy T, Reddy SP, Yamamoto M, Kensler TW, and Biswal S (2006a) Nrf2 is a critical regulator of the innate immune response and survival during experimental sepsis. J Clin Invest 116:984–995.
Thimmulappa RK, Scollick C, Traore K, Yates M, Trush MA, Liby KT, Sporn MB, Yamamoto M, Kensler TW, and Biswal S (2006b) Nrf2-dependent protection from LPS induced inflammatory response and mortality by CDDO-Imidazolide. Biochem Biophys Res Commun 351:883–889.
Thomson L, Tenopoulou M, Lightfoot R, Tsika E, Parastatidis I, Martinez M, Greco TM, Doulias PT, Wu Y, Tang WH, et al. (2012) Immunoglobulins against tyrosinenitrated epitopes in coronary artery disease. Circulation 126:2392–2401.
Todorovic M, Newman JR, Shan J, Bentley S, Wood SA, Silburn PA, and Mellick GD (2015) Comprehensive assessment of genetic sequence variants in the antioxidant ‘master regulator’ NRF2 in idiopathic Parkinson’s disease. PLoS One 10:e0128030.
Tong KI, Padmanabhan B, Kobayashi A, Shang C, Hirotsu Y, Yokoyama S, and Yamamoto M (2007) Different electrostatic potentials define ETGE and DLG motifs as hinge and latch in oxidative stress response. Mol Cell Biol 27:7511–7521.
Tsai HH, Lai HY, Chen YC, Li CF, Huang HS, Liu HS, Tsai YS, and Wang JM (2017) Metformin promotes apoptosis in hepatocellular carcinoma through the CEBPDinduced autophagy pathway. Oncotarget 8:13832–13845.
Tsuchida K, Tsujita T, Hayashi M, Ojima A, Keleku-Lukwete N, Katsuoka F, Otsuki A, Kikuchi H, Oshima Y, Suzuki M, et al. (2017) Halofuginone enhances the chemosensitivity of cancer cells by suppressing NRF2 accumulation. Free Radic Biol Med 103:236–247.
Tu J, Zhang X, Zhu Y, Dai Y, Li N, Yang F, Zhang Q, Brann DW, and Wang R (2015) Cell-permeable peptide targeting the Nrf2-Keap1 interaction: a potential novel therapy for global cerebral ischemia. J Neurosci 35:14727–14739.
Tung MC, Lin PL, Wang YC, He TY, Lee MC, Yeh SD, Chen CY, and Lee H (2015) Mutant p53 confers chemoresistance in non-small cell lung cancer by upregulating Nrf2. Oncotarget 6:41692–41705.
Türei D, Papp D, Fazekas D, Földvári-Nagy L, Módos D, Lenti K, Csermely P, and Korcsmáros T (2013) NRF2-ome: an integrated web resource to discover protein interaction and regulatory networks of NRF2. Oxid Med Cell Longev 2013: 737591.
Tzima S, Victoratos P, Kranidioti K, Alexiou M, and Kollias G (2009) Myeloid heme oxygenase-1 regulates innate immunity and autoimmunity by modulating IFNbeta production. J Exp Med 206:1167–1179.
Ungvari Z, Bagi Z, Feher A, Recchia FA, Sonntag WE, Pearson K, de Cabo R, and Csiszar A (2010) Resveratrol confers endothelial protection via activation of the antioxidant transcription factor Nrf2. Am J Physiol Heart Circ Physiol 299: H18–H24.
Ungvari Z, Bailey-Downs L, Gautam T, Jimenez R, Losonczy G, Zhang C, Ballabh P, Recchia FA, Wilkerson DC, Sonntag WE, et al. (2011) Adaptive induction of NF-E2- related factor-2-driven antioxidant genes in endothelial cells in response to hyperglycemia. Am J Physiol Heart Circ Physiol 300:H1133–H1140.
Uruno A, Furusawa Y, Yagishita Y, Fukutomi T, Muramatsu H, Negishi T, Sugawara A, Kensler TW, and Yamamoto M (2013) The Keap1-Nrf2 system prevents onset of diabetes mellitus. Mol Cell Biol 33:2996–3010.
Uruno A, Yagishita Y, and Yamamoto M (2015) The Keap1-Nrf2 system and diabetes mellitus. Arch Biochem Biophys 566:76–84.
van Horssen J, Drexhage JA, Flor T, Gerritsen W, van der Valk P, and de Vries HE (2010) Nrf2 and DJ1 are consistently upregulated in inflammatory multiple sclerosis lesions. Free Radic Biol Med 49:1283–1289.
van Muiswinkel FL, de Vos RA, Bol JG, Andringa G, Jansen Steur EN, Ross D, Siegel D, and Drukarch B (2004) Expression of NAD(P)H:quinone oxidoreductase in the normal and Parkinsonian substantia nigra. Neurobiol Aging 25:1253–1262.
Vidal M, Cusick ME, and Barabási AL (2011) Interactome networks and human disease. Cell 144:986–998.
von Otter M, Bergström P, Quattrone A, De Marco EV, Annesi G, Söderkvist P, Wettinger SB, Drozdzik M, Bialecka M, Nissbrandt H, et al. (2014) Genetic associations of Nrf2-encoding NFE2L2 variants with Parkinson’s disease: a multicenter study. BMC Med Genet 15:131.
von Otter M, Landgren S, Nilsson S, Celojevic D, Bergström P, Håkansson A, Nissbrandt H, Drozdzik M, Bialecka M, Kurzawski M, et al. (2010a) Association of Nrf2-encoding NFE2L2 haplotypes with Parkinson’s disease. BMC Med Genet 11: 36.
von Otter M, Landgren S, Nilsson S, Zetterberg M, Celojevic D, Bergström P, Minthon L, Bogdanovic N, Andreasen N, Gustafson DR, et al. (2010b) Nrf2-encoding NFE2L2 haplotypes influence disease progression but not risk in Alzheimer’s disease and age-related cataract. Mech Ageing Dev 131:105–110.
Wang L, Han J, Shan P, You S, Chen X, Jin Y, Wang J, Huang W, Wang Y, and Liang G (2017a) MD2 blockage protects obesity-induced vascular remodeling via activating AMPK/Nrf2. Obesity (Silver Spring) 25:1532–1539.
Wang R, An J, Ji F, Jiao H, Sun H, and Zhou D (2008) Hypermethylation of the Keap1 gene in human lung cancer cell lines and lung cancer tissues. Biochem Biophys Res Commun 373:151–154.
Wang X, Chen H, Liu J, Ouyang Y, Wang D, Bao W, and Liu L (2015) Association between the NF-E2 related factor 2 gene polymorphism and oxidative stress, antioxidative status, and newly-diagnosed type 2 diabetes mellitus in a Chinese population. Int J Mol Sci 16:16483–16496.
Wang X, de Rivero Vaccari JP, Wang H, Diaz P, German R, Marcillo AE, and Keane RW (2012) Activation of the nuclear factor E2-related factor 2/antioxidant response element pathway is neuroprotective after spinal cord injury. J Neurotrauma 29: 936–945.
Wang X, Hausding M, Weng SY, Kim YO, Steven S, Klein T, Daiber A, and Schuppan D (2018) Gliptins suppress inflammatory macrophage activation to mitigate inflammation, fibrosis, oxidative stress, and vascular dysfunction in models of nonalcoholic steatohepatitis and liver fibrosis. Antioxid Redox Signal. 28:87–109.
Wang X, Li M, Cao Y, Wang J, Zhang H, Zhou X, Li Q, and Wang L (2017b) Tenuigenin inhibits LPS-induced inflammatory responses in microglia via activating the Nrf2-mediated HO-1 signaling pathway. Eur J Pharmacol 809:196–202.
Wang XJ, Hayes JD, Henderson CJ, and Wolf CR (2007) Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha. Proc Natl Acad Sci USA 104:19589–19594.
Wang Y, Huang Y, Xu Y, Ruan W, Wang H, Zhang Y, Saavedra JM, Zhang L, Huang Z, and Pang T (2017c) A dual AMPK/Nrf2 activator reduces brain inflammation after stroke by enhancing microglia M2 polarization. Antioxid Redox Signal 28: 141–163.
Wasik U, Milkiewicz M, Kempinska-Podhorodecka A, and Milkiewicz P (2017) Protection against oxidative stress mediated by the Nrf2/Keap1 axis is impaired in primary biliary cholangitis. Sci Rep 7:44769.
Wen X, Thorne G, Hu L, Joy MS, and Aleksunes LM (2015) Activation of NRF2 signaling in HEK293 cells by a first-in-class direct KEAP1-NRF2 inhibitor. J Biochem Mol Toxicol 29:261–266.
Wenzel P, Kossmann S, Münzel T, and Daiber A (2017) Redox regulation of cardiovascular inflammation: immunomodulatory function of mitochondrial and Noxderived reactive oxygen and nitrogen species. Free Radic Biol Med 109:48–60.
Wenzel P, Oelze M, Coldewey M, Hortmann M, Seeling A, Hink U, Mollnau H, Stalleicken D, Weiner H, Lehmann J, et al. (2007) Heme oxygenase-1: a novel key player in the development of tolerance in response to organic nitrates. Arterioscler Thromb Vasc Biol 27:1729–1735.
Wenzel P, Rossmann H, Müller C, Kossmann S, Oelze M, Schulz A, Arnold N, Simsek C, Lagrange J, Klemz R, et al. (2015) Heme oxygenase-1 suppresses a proinflammatory phenotype in monocytes and determines endothelial function and arterial hypertension in mice and humans. Eur Heart J 36:3437–3446.
Winkel AF, Engel CK, Margerie D, Kannt A, Szillat H, Glombik H, Kallus C, Ruf S, Güssregen S, Riedel J, et al. (2015) Characterization of RA839, a noncovalent small molecule binder to Keap1 and selective activator of Nrf2 signaling. J Biol Chem 290:28446–28455.
Winston JT, Strack P, Beer-Romero P, Chu CY, Elledge SJ, and Harper JW (1999) The SCFbeta-TRCP-ubiquitin ligase complex associates specifically with phosphorylated destruction motifs in IkappaBalpha and beta-catenin and stimulates IkappaBalpha ubiquitination in vitro. Genes Dev 13:270–283.
Wu Q, Wang Q, Mao G, Dowling CA, Lundy SK, and Mao-Draayer Y (2017) Dimethyl fumarate selectively reduces memory T cells and shifts the balance between Th1/ Th17 and Th2 in multiple sclerosis patients. J Immunol 198:3069–3080.
Wu W, Zhao L, Yang P, Zhou W, Li B, Moorhead JF, Varghese Z, Ruan XZ, and Chen Y (2016a) Inflammatory stress sensitizes the liver to atorvastatin-induced injury in ApoE-/- mice. PLoS One 11:e0159512.
Wu WJ, Jia WW, Liu XH, Pan LL, Zhang QY, Yang D, Shen XY, Liu L, and Zhu YZ (2016b) S-propargyl-cysteine attenuates inflammatory response in rheumatoid arthritis by modulating the Nrf2-ARE signaling pathway. Redox Biol 10:157–167.
Xia N, Daiber A, Förstermann U, and Li H (2017) Antioxidant effects of resveratrol in the cardiovascular system. Br J Pharmacol 174:1633–1646.
Xiaobo C, Majidi M, Feng M, Shao R, Wang J, Zhao Y, Baladandayuthapani V, Song J, Fang B, Ji L, et al. (2016) TUSC2(FUS1)-erlotinib induced vulnerabilities in epidermal growth factor receptor (EGFR) wildtype non-small cell lung cancer (NSCLC) targeted by the repurposed drug auranofin. Sci Rep 6:35741.
Xie H, Zhou F, Liu L, Zhu G, Li Q, Li C, and Gao T (2016) Vitiligo: how do oxidative stress-induced autoantigens trigger autoimmunity? J Dermatol Sci 81:3–9.
Xiong M, Li B, Zhu Q, Wang YX, and Zhang HY (2014) Identification of transcription factors for drug-associated gene modules and biomedical implications. Bioinformatics 30:305–309.
Xu IM, Lai RK, Lin SH, Tse AP, Chiu DK, Koh HY, Law CT, Wong CM, Cai Z, Wong CC, et al. (2016a) Transketolase counteracts oxidative stress to drive cancer development. Proc Natl Acad Sci USA 113:E725–E734.
Xu J, Donepudi AC, Moscovitz JE, and Slitt AL (2013) Keap1-knockdown decreases fasting-induced fatty liver via altered lipid metabolism and decreased fatty acid mobilization from adipose tissue. PLoS One 8:e79841.
Xu X, Sun J, Chang X, Wang J, Luo M, Wintergerst KA, Miao L, and Cai L (2016b) Genetic variants of nuclear factor erythroid-derived 2-like 2 associated with the complications in Han descents with type 2 diabetes mellitus of Northeast China. J Cell Mol Med 20:2078–2088.
Xu Z, Wei Y, Gong J, Cho H, Park JK, Sung ER, Huang H, Wu L, Eberhart C, Handa JT, et al. (2014) NRF2 plays a protective role in diabetic retinopathy in mice. Diabetologia 57:204–213.
Xu Z, Zhang F, Sun F, Gu K, Dong S, and He D (2015) Dimethyl fumarate for multiple sclerosis. Cochrane Database Syst Rev 4:CD011076.
Xue P, Hou Y, Chen Y, Yang B, Fu J, Zheng H, Yarborough K, Woods CG, Liu D, Yamamoto M, et al. (2013) Adipose deficiency of Nrf2 in ob/ob mice results in severe metabolic syndrome. Diabetes 62:845–854.
Yamamoto T, Suzuki T, Kobayashi A, Wakabayashi J, Maher J, Motohashi H, and Yamamoto M (2008) Physiological significance of reactive cysteine residues of Keap1 in determining Nrf2 activity. Mol Cell Biol 28:2758–2770.
Yamamoto T, Yoh K, Kobayashi A, Ishii Y, Kure S, Koyama A, Sakamoto T, Sekizawa K, Motohashi H, and Yamamoto M (2004) Identification of polymorphisms in the promoter region of the human NRF2 gene. Biochem Biophys Res Commun 321: 72–79.
Yang M, Yao Y, Eades G, Zhang Y, and Zhou Q (2011) MiR-28 regulates Nrf2 expression through a Keap1-independent mechanism. Breast Cancer Res Treat 129: 983–991.
Yao W, Zhang JC, Ishima T, Dong C, Yang C, Ren Q, Ma M, Han M, Wu J, Suganuma H, et al. (2016) Role of Keap1-Nrf2 signaling in depression and dietary intake of glucoraphanin confers stress resilience in mice. Sci Rep 6:30659.
Yap JL, Worlikar S, MacKerell AD Jr, Shapiro P, and Fletcher S (2011) Smallmolecule inhibitors of the ERK signaling pathway: towards novel anticancer therapeutics. ChemMedChem 6:38–48.
Yoh K, Hirayama A, Ishizaki K, Yamada A, Takeuchi M, Yamagishi S, Morito N, Nakano T, Ojima M, Shimohata H, et al. (2008) Hyperglycemia induces oxidative and nitrosative stress and increases renal functional impairment in Nrf2-deficient mice. Genes Cells 13:1159–1170.
Yore MM, Kettenbach AN, Sporn MB, Gerber SA, and Liby KT (2011) Proteomic analysis shows synthetic oleanane triterpenoid binds to mTOR. PLoS One 6: e22862.
Yoshizaki Y, Mori T, Ishigami-Yuasa M, Kikuchi E, Takahashi D, Zeniya M, Nomura N, Mori Y, Araki Y, Ando F, et al. (2017) Drug-repositioning screening for Keap1- Nrf2 binding inhibitors using fluorescence correlation spectroscopy. Sci Rep 7: 3945.
Yu M, Li H, Liu Q, Liu F, Tang L, Li C, Yuan Y, Zhan Y, Xu W, Li W, et al. (2011) Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell Signal 23:883–892.
Zehir A, Benayed R, Shah RH, Syed A, Middha S, Kim HR, Srinivasan P, Gao J, Chakravarty D, Devlin SM, et al. (2017) Mutational landscape of metastatic cancer revealed from prospective clinical sequencing of 10,000 patients. Nat Med 23: 703–713.
Zeidan TA, Duncan S, Hencken CP, Wynn TA, and Sanrame CN (2014) Prodrugs of Fumarates and Their Use in Treating Various Diseases, Alkermes Pharma Ireland, Dublin, Ireland. Zhang DD (2013) Bardoxolone brings Nrf2-based therapies to light. Antioxid Redox Signal 19:517–518.
Zhang QY, Chu XY, Jiang LH, Liu MY, Mei ZL, and Zhang HY (2017) Identification of non-electrophilic Nrf2 activators from approved drugs. Molecules 22:1–12.
Zhao C, Zhang Y, Liu H, Li P, Zhang H, and Cheng G (2017) Fortunellin protects against high fructose-induced diabetic heart injury in mice by suppressing inflammation and oxidative stress via AMPK/Nrf-2 pathway regulation. Biochem Biophys Res Commun 490:552–559.
Zhao M, Chen H, Ding Q, Xu X, Yu B, and Huang Z (2016) Nuclear factor erythroid 2-related factor 2 deficiency exacerbates lupus nephritis in B6/lpr mice by regulating Th17 cell function. Sci Rep 6:38619.
Zhou X, Menche J, Barabási AL, and Sharma A (2014) Human symptoms-disease network. Nat Commun 5:4212.
本文由福山生物整理翻译,转载请注明出处。
目 录
缩写:
AD: 阿尔茨海默病; ALS: 肌萎缩侧索硬化症; AMPK,AMP活化蛋白激酶; ARE: 抗氧化反应元件; β-TrCP: 含有E3泛素蛋白连接酶的β-转导蛋白重复序列; bZip: 碱性区域 - 亮氨酸拉链; CDDO: 2-氰基-3,12-二氧代-油酸-1,9(11)- 二烯-28-酸; CDDO-Me: CDDO-甲酯; COPD:慢性阻塞性肺病; CUL3,Cullin 3; DC:树突状细胞; DMF: 富马酸二甲酯; EAE:实验性自身免疫性脑脊髓炎; ECH:具有Cap'n'collar同源性的红系细胞衍生蛋白; GCLC:g-谷氨酰半胱氨酸连接酶催化亚基; GCLM:g-谷氨酰半胱氨酸连接酶调节剂亚基; GI:胃肠道; GSH:谷胱甘肽; GSK-3:糖原合成酶激酶3; HFD,高脂饮食; HNSCC:头颈部鳞状细胞癌; HO-1,血红素加氧酶-1; IBD:炎症性肠病; IκB:κ-B抑制剂; IKK:IκB激酶; IL:白细胞介素; KEAP1:kelch样ECH相关蛋白1; LDL:低密度脂蛋白; LPS:脂多糖; MAF:肌肉腱膜纤维肉瘤蛋白; MMF:富马酸单甲酯; MS:多发性硬化; NASH:非酒精性脂肪性肝炎; NF-κB:活化B细胞核因子κ-轻链增强子的p65亚基; NQO1:NADPH:醌氧化还原酶; NRF2:核因子(红细胞衍生的2)样2; PD:帕金森病; PI3K:磷脂酰肌醇3-激酶; PPI:蛋白质-蛋白质相互作用; PTEN:磷酸酶和张力蛋白同系物; RA: 类风湿性关节炎; RBX1:RING-box蛋白1; RNS:活性氮物种; ROS:活性氧; SFN:萝卜硫素; SLE:系统性红斑狼疮; SNP:单核苷酸多态性; SQSTM1:sequestosome 1; STAT:信号转导和转录激活因子; T2DM:2型糖尿病; TGF:转化生长因子; Th,T辅助子; TNF:肿瘤坏死因子; Treg:T调控。
运用系统医学方法阐明转录因子NRF2作为慢性疾病的治疗靶标
Antonio Cuadrado et al., PHARMACOLOGICAL REVIEWS, 2018
摘要
系统医学是基于疾病机理,而非基于具体症状或器官的疾病研究方法,并以非假设驱动的方式识别治疗靶标。本论文中,我们将系统医学方法应用于转录因子核因子( NRF2)的研究,通过交叉验证其在蛋白质-蛋白质相互作用网络(NRF2相互作用组)中的作用,该相互作用网络在功能上与低水平应激、慢性炎症、代谢改变和活性氧形成有关的细胞保护相关联。这些分子谱的多尺度网络分析表明,NRF2表达和活性的改变是疾病子网路中的常见机制(NRF2病变)。该相互作用网络关联了明显的异质性表型,如自身免疫、呼吸、消化、心血管、代谢和神经退行性疾病,以及癌症。重要的是,系统医学方法在计算机上匹配并证实了在临床开发的不同阶段在体内验证的NRF2调节药物的若干应用。从药理学上看,这些药物的特征多样,有亲电的富马酸二甲酯、合成的三萜类化合物如巴多索酮甲基和萝卜硫素、蛋白-蛋白或DNA -蛋白相互作用抑制剂,甚至有注册的药物如二甲双胍和他汀类药物,它们可以激活NRF2并且可能被重新用于指示 NRF2疾病表型群。因此,NRF2代表了经典和系统医学方法充分接受的第一类目标之一,这些方法通过关注一组似乎具有机械联系的疾病表型来促进药物开发和药物新用。 因此,由此产生的NRF2药物组可能会迅速为这类慢性疾病提供一些令人惊讶的临床选择。
Ⅰ. 引言
在上个世纪,寿命几乎翻了一番,特定老龄化疾病现在变得普遍。然而,大多数疾病的病理机制很难理解,治疗方法也是通过纠正症状或危险因素来治疗。此外,与迄今为止只考虑一种疾病、一种药物的线性方法相反,慢性病显示出高度的关联性,需要更精确的、以机制为基础的疾病定义,而不是目前以器官和症状为基础的疾病定义。在人类基因组测序和分子网络的发展之后,一种新的疾病概念由此产生,疾病的诊断不仅是通过临床症状,而且主要是通过潜在的分子特征识别(Goh et al., 2007)。不同的病理表型具有共同的分子机制,这一事实也为总结为“多种疾病,一种药物”和药物新用的治疗新概念提供了理论基础。网络医学,即将网络概念应用于疾病和药物之间的动态联系分析,为开发这种新方法提供了新机会。老年人慢性病很可能的特征是衰老过程中体内平衡的丧失或环境因素的影响,所有这些都会通过病理性形成活性氧(ROS),慢性炎症和代谢失调导致低水平应激。基于网络医学方法,我们将在本文中提供大量证据,表明核因子(erythroid-derived 2)–like 2 (NRF2)作为多种细胞保护反应的主调控因子和特定疾病集群中的一个关键分子节点,为药物开发和药物新用提供了一种新的策略。Ⅱ. 从NRF2相互作用组到NRF2疾病网络
A. NRF2作为一个细胞稳态的主要调节器
NRF2是一种碱性区域亮氨酸拉链(bZip)转录因子(图1),在细胞核内与小肌筋膜纤维瘤蛋白(MAF) K、G、F形成异二聚体。异二聚体识别一个称为抗氧化应答元件(ARE)的增强子序列,其存在于超过250个基因(ARE基因)的调节区域中(Ma,2013;Hayes and Dinkova-Kostova,2014)。这些基因编码参与I期、II期和III期生物转化反应和抗氧化机制的合作酶网络,产生NADPH、谷胱甘肽(GSH)和硫氧还蛋白反应;脂质和铁的分解代谢;以及与其他转录因子的相互作用等(Hayes and Dinkova-Kostova, 2014)。最近,NRF2也被发现可调控多个蛋白酶体亚基和自噬基因的表达,为其调控蛋白酶体平衡提供了额外的研究兴趣(Pajares et al., 2015, 2016, 2017;de la Vega等,2016)。 NRF2在临床上的重要意义在于它可能具有药理学靶向性,对患者有益。调节NRF2转录活性的主要机制是通过E3连接酶适配器KEAP1(具有Cap'n'collar同源性(ECH)的Kelch样红系细胞衍生蛋白 - 相关蛋白1)来控制蛋白的稳定(图2)。KEAP1是同型二聚体蛋白,其将NRF2与Cullin 3和RING-box蛋白1(CUL3 / RBX1)形成的E3连接酶复合物连接。在稳态条件下,KEAP1同二聚体的N-末端结构域在两个氨基酸序列上与一个分子的NRF2结合,具有低(天冬氨酸,亮氨酸和甘氨酸; DLG)和高(谷氨酸,苏氨酸,甘氨酸和谷氨酸; ETGE)亲和力,因此,通过CUL3 / RBX1(Tong等人,2007)将NRF2呈现为泛素化,并随后通过蛋白酶体降解。KEAP1是一种氧化还原和亲电子传感器,在关键半胱氨酸修饰后失去其抑制NRF2的能力(图2;生物标志物作为NRF2特征并参与监测目标)。调节NRF2稳定性的另一种机制是由糖原合成酶激酶3(GSK-3)介导的磷酸化(图2)。这种激酶磷酸化NRF2(天冬氨酸,丝氨酸,甘氨酸,异亮氨酸,丝氨酸; DSGIS)的结构域,因此为含有E3泛素蛋白连接酶(b-TrCP)的E3连接酶接头b-转导蛋白重复序列产生识别基序,该基序将NRF2呈递给 CUL1 / RBX1复合物,导致NRF2泛素依赖性蛋白酶体降解的替代途径。因此,KEAP1和GSK-3 / b-TrCP分别在氧化还原稳态和细胞信号传导的情况下严格控制NRF2蛋白水平(Cuadrado,2015)。已经报道了NRF2在蛋白质,mRNA或基因水平上调节的其他机制(Hayes and Dinkova-Kostova,2014),但至少这两种机制适合于药理学调节。
图2. KEAP1和β-TrCP对NRF2稳定性的调节及其药理学靶向。(A)根据双重调节模型(Rada等,2011),NRF2的两个结构域,称为Neh2和Neh6,参与NRF2降解分别响应氧化还原和亲电子变化(KEAP1)和信号激酶(β-TrCP)。Neh2结构域结合E3连接酶适配子KEAP1,其呈递NRF2用于泛素化至CUL3 / RBX1复合物。 Neh6结构域需要GSK-3的先前磷酸化以结合E3连接酶适配子β-TrCP并随后通过CUL1/ RBX1复合物进行泛素化(详见文本)。(B)NRF2和KEAP1之间结合的细节以及当前针对这种相互作用的策略。KEAP1同型二聚体在Neh2结构域的两个基序处结合NRF2:低亲和力(29-DLG-31)和高亲和力(79-ETGE-82)结合位点。目前破坏这种相互作用的策略包括:改变半胱氨酸C151,273和288的巯基的亲电试剂; 改变NRF2与KEAP1的DC结构域的对接的PPI抑制剂。(C)NRF2和β-TrCP的假设结合,并提出了针对这种相互作用的策略。当它被GSK-3(Rada等,2011,2012)磷酸化时,β-TrCP同型二聚体在磷酸基序334-DSGIS-338处与Nhe6结构域结合,当与磷酸基序373-DSAPGS-378结合时独立于GSK-3(Chowdhry等,2013)。在该图中,我们假设,与KEAP1类似,一个β-TrCP同二聚体在两个磷酸基序上与一个NRF2分子相互作用,但仍缺乏实验证据。破坏这种相互作用的两种可能策略包括使用GSK-3抑制剂和PPI抑制剂。
B. NRF2及其调控途径在人类相互作用组和疾病组的定位
编码NRF2的基因,称为NFE2L2,具有高度多态性,诱变频率为每72 bp 1次。关于该主题的优秀综述在2015年报告了多达18个单核苷酸多态性(SNPs),其中大多数在5’调控区域和内含子1中(Cho等人,2015)。这些SNPs中的一些可能构成与慢性疾病发作或进展风险相关的功能性单倍型。功能性单倍型的变异可能对表现出特定疾病的临床症状的个体的比例具有微妙的影响,但是它们可能在群体水平上具有深远的影响,并且可能定义在精准医疗中靶向该基因的具体策略。网络医学的最新进展提供了定量工具来表征基因与其相互作用(相互作用组)之间的相互作用如何与病理学相关(Barabasi等,2011; Vidal等,2011; Guney等,2016),分子网络失调在各种疾病中如何普遍(Menche等,2015),以及疾病如何在特定组织中表现(Kitsak等,2016)。为了从系统医学的角度理解NRF2在病理学中的相关性,首先我们已经生成了人类相互作用组图。我们整合并策划了与NRF2调节途径相关的蛋白质之间的物理相互作用的信息(Hayes and Dinkova-Kostova,2014; Cuadrado,2015)。交互数据来自最近发表的人类相互作用组,其汇编了几种蛋白质 - 蛋白质相互作用(PPI)资源的数据(Turei等,2013; Menche等,2015)。然而,目前可用于NRF2相互作用组开发的信息受限于以下事实:因为NRF2是非常短的半衰期蛋白质,所以可能未检测到一些有意义的相互作用。然而,在相互作用组中发现了一些与NRF2物理性相互作用的众所周知的蛋白质,包括KEAP1,b-TrCP和MAF。另一组NRF2相互作用蛋白对应于具有调节基因表达功能的核蛋白。这些包括与bZip转录因子,核受体或共激活因子相关的蛋白质,或涉及组蛋白乙酰化的蛋白质。因此,NRF2相互作用组证明除了与该转录因子直接相关的基因调控之外的其他基因调控机制。NRF2在几个残基处被磷酸化,因此预期它与几种激酶相互作用,所述激酶包括GSK-3和几种蛋白激酶C同种型。这些激酶是一些膜受体和衔接子/支架蛋白的下游。除了这种物理相互作用,我们已经确定了NRF2邻域中富集的几种生物功能,包括代谢过程,如戊糖,四吡咯,血红素,葡萄糖6-磷酸,半胱氨酸,GSH,甘油醛-3-磷酸和NADPH的生物合成。大多数这些调节蛋白不直接相互作用,而是通过作为介质的蛋白质连接(图3;特定相互作用分子的更详细描述可在这个网址找到http://sbi.imim.es/data/nrf2/)。
已经报道了几种疾病中NRF2相互作用组的扰动。我们基于DisGeNET (Pinero et al., 2017)和GeneCards (Stelzer et al., 2016)数据库,结合部分动物模型研究的知识,整理了37种nrf2相关疾病的列表,绘制了疾病组图。我们还使用DisGeNET,OMIM和GWAS数据库检索了这些病症的疾病 - 基因关联(Menche等,2015)(表1)。NRF2与每种NRF2相关疾病表型的已知疾病基因的基于相互作用组的接近度(Guney和Oliva,2014)显示在图4中。与随机选择的蛋白质相比,NRF2与消化系统和癌症(如前列腺,肝脏和肺肿瘤)的已知疾病基因显着接近,突出了NRF2在这些病理中的关键功能作用。此外,发现NRF2与代谢和心血管疾病相关的各种蛋白质均有近端表达,例如糖尿病,高血糖症,局部缺血,大脑中动脉梗塞和动脉粥样硬化。NRF2的蛋白质相互作用还将其与呼吸系统疾病相关的基因(如哮喘,肺纤维化和肺气肿)以及神经退行性疾病(如阿尔茨海默病(AD),帕金森病(PD)和肌萎缩侧索硬化症(ALS))联系起来。
表1.有NRF2相关证据的疾病群集
从DisGeNet数据库中选择具有与NRF2遗传相关的证据的疾病表型。DisGeNet整合来自各种资源的疾病基因关联信息,如UniProt,ClinVar,GWAS目录和比较毒物基因组学数据库,并根据资源数量和支持这些关联的出版物评估疾病基因关联。该清单的制定基于以下标准:1)仅选择在Pubmed中具有多于一个引用的病理表型; 2)可靠性评分设定为0.001的阈值; 3)具有非常相似的名称或重叠术语的疾病条目被简化为单个条目。
| 病理表型 | 可靠性分数 | 病理表型 | 可靠性分数 |
| 糖尿病肾病 | 0.2016 | 糖尿病性心肌病 | 0.0803 |
| 肝硬化 | 0.2005 | 大脑中动脉梗塞 | 0.0800 |
| 非酒精性脂肪性肝炎 | 0.2005 | 乳腺肿瘤 | 0.0087 |
| 急性肾损伤 | 0.2000 | 白癜风 | 0.0076 |
| 肺纤维化 | 0.2000 | 动脉粥样硬化 | 0.0067 |
| 非小细胞肺癌 | 0.1252 | 哮喘 | 0.0043 |
| 鳞状细胞癌 | 0.1243 | 白血病 | 0.0038 |
| 肝肿瘤 | 0.1238 | 结肠肿瘤 | 0.0038 |
| 高血糖 | 0.1208 | 胃肠疾病 | 0.0029 |
| 药物性肝损伤 | 0.1200 | 帕金森综合症 | 0.0026 |
| 前列腺肿瘤 | 0.1200 | 系统性红斑狼疮性肾炎 | 0.0026 |
| 慢性阻塞性肺疾病 | 0.0899 | 胶质瘤 | 0.0024 |
| 结直肠肿瘤 | 0.0847 | 肌萎缩侧索硬化症 | 0.0022 |
| 阿尔茨海默病 | 0.0837 | 缺血 | 0.0016 |
| 2型糖尿病 | 0.0814 | 肺气肿 | 0.0013 |
| 慢性肾病 | 0.0808 | 胰腺肿瘤 | 0.0013 |
| 糖尿病性视网膜病变 | 0.0805 | 血管疾病 | 0.0013 |
| 亨廷顿氏病 | 0.0805 | 脓血症 | 0.0013 |
因此,基于相互作用组的接近度提供了关于NRF2如何与各种病理状况相关联的观点。之前已将疾病之间的关系概括为一种网络,成为疾病组,其基于遗传(Goh等人,2007)和临床(Hidalgo等人,2009; Zhou等人,2014)的共同性来连接它们。在图5中,我们基于共享基因,症状相似性和合并症定义了疾病网络,即NRF2疾病组。NRF2似乎连接基本上由炎症过程控制的疾病,例如急性肾损伤,肝硬化和动脉粥样硬化。此外,AD,PD,亨廷顿舞蹈病和ALS等神经退行性疾病构成一个集群,与最近的研究一致,也暗示NRF2在神经炎症过程中的作用(Rojo等,2010; Lastres-Becker等,2012,2014; Jung等,2017; Wang等,2017b)。通过激酶信号转导级联对NRF2的调控(Jung et al., 2017)可以解释密切相关的癌症簇,特别是支持NRF2参与结肠和乳腺肿瘤病理性ROS形成(Gonzalez-Donquiles et al., 2017;Lu et al., 2017)。
Ⅲ. NRF2在人类疾病状态中的靶标验证
A. NRF2在炎症消退中的关键作用
持续性炎症是NRF2疾病组中发现的所有病理表型的标志。这很可能是因为炎症与活性氧(ROS)的局部和全身病理形成增加有关。事实上,ROS和活性氮物种(RNS)刺激和加重炎症反应,这些反应与转录因子NF-κB的激活机制相关的激活机制相关(NF-κB:激活的B细胞的核因子κ-轻链增强子的p65亚基)(Wenzel等,2017)。非常简化的,在静息免疫细胞中,NF-κB通过与核κ-B抑制因子(IκBα)的相互作用保留在细胞溶质中。源自微生物的病原体相关分子模式分子以及响应于组织损伤而释放的损伤相关分子模式分子刺激免疫细胞表达的同源受体,其导致IκB激酶(IKK)β的激活。该激酶使IκBα磷酸化,使其靶向降解并允许核转位和NF-κB活化(Napetschnig和Wu,2013)。这些事件通过IκBα的几种调节模式进行氧化还原控制(Bowie和O'Neill,2000; Morgan和Liu,2011; Siomek,2012),但最近描述的一种模式涉及KEAP1对IKKβ稳定性的调节。就像NRF2一样,IKKβ具有ETGE基序,使其能够与KEAP结合,进行泛素化和蛋白酶体降解。因此,在基础氧化还原条件下,活性KEAP1靶向IKKβ进行降解,然后IκBα抑制NF-κB。相反,在存在ROS的情况下,KEAP1被抑制并且IKKβ稳定,磷酸化IκBα并导致其降解并因此导致NF-κB的上调(Lee等人,2009)。由于NRF2是氧化还原稳态的主要调节因子,因此它对NF-κB活性进行间接控制。脂多糖(LPS)同时激活快速、促炎性的NF-κB反应和缓慢的NRF2反应。当NRF2最大活性时,NF-κB响应随后被抑制(Cuadrado等,2014)。例如,Ras相关的C3肉毒杆菌毒素底物1(Rho家族的一个小G蛋白)激活NF-kB途径,并且NRF2过表达被阻断,而NRF2敲低增强NF-κB依赖性转录(Cuadrado等,2014)。一致地,在用LPS或肿瘤坏死因子(TNF)-α攻击的NRF2缺陷型(Nrf2-/-)小鼠中,IKK的活性恶化并导致IκB的磷酸化和降解增加(Thimmulappa等,2006a)。
NRF2还诱导抗炎表型,其调节CD8+T细胞(Sha等人,2015)以及巨噬细胞和小胶质细胞的功能(Rojo等人,2010,2014a; Brune等人,2013)。这是因为NRF2通过调节胱氨酸/谷氨酸转运蛋白和GSH合成酶?-谷氨酰半胱氨酸连接酶调节器和催化亚基[?-谷氨酰半胱氨酸连接酶调节剂亚基(GCLM)和?-谷氨酰半胱氨酸连接酶催化亚基(GCLC)]来增加巨噬细胞中的半胱氨酸和GSH水平。相反,GSH耗竭使巨噬细胞对LPS激活NRF2敏感(Diotallevi等,2017)。所有这些研究都指出NRF2是一种抗炎因子,对控制炎症反应的强度和持续时间至关重要(图6)。
通过前馈和反馈机制产生NRF2和NF-κB串扰(图7)。在转录水平,由于NFE2L2基因的启动子区域中存在几个功能性结合位点,NF-κB激活NRF2表达,从而诱导负反馈环(Rushworth等,2012)。此外,NF-κB和NRF2转录因子都需要共激活因子CBP / p300,它是组蛋白乙酰转移酶,乙酰化并增加DNA结合能力。因此,NF-κB过表达阻碍了NRF2的CBP / p300的可用性,因此降低了其转录能力,而NF-kB敲低显示出相反的效果(Liu等人,2008)。另外,NF-κB可以促进组蛋白脱乙酰酶-3与MAF蛋白的相互作用,因此阻止它们与NRF2的二聚化(Liu等人,2008)。NF-κB将KEAP1结合并转移至细胞核,从而有利于NRF2在该细胞区室中的泛素化和降解(Yu等,2011)。 E3连接酶适配子β-TrCP标记IκBα(Winston等人,1999)和NRF2(Rada等人,2011,2012; Cuadrado,2015)用于蛋白酶体降解,因此它可以导致增加的NF-kB活性。
NRF2的抗炎活性被认为仅依赖于氧化还原代谢的调节或与NF-κB的串扰。然而,NRF2还可以在暴露于LPS后直接阻断巨噬细胞中促炎基因白细胞介素(IL)-6和IL-1β的转录(Kobayashi等,2016)。LPS暴露或NRF2的药理学活化导致其与这些促炎基因的近端启动子结合并阻断RNA pol II的募集。该机制似乎与NRF2与其成熟的ARE增强子的结合无关。在其他研究中,NRF2可以直接调节其他几种巨噬细胞特异性基因的表达,例如具有胶原结构的巨噬细胞受体,细菌吞噬作用所需的受体,或CD36,氧化低密度脂蛋白的清道夫受体(Harvey等,2011;Ishii和Mann,2014)。类似地,编码促炎细胞因子IL-17D的基因含有AREs,并且该NRF2-T辅助因子(Th)17轴似乎赋予针对肿瘤发生和病毒感染的保护作用(Saddawi-Konefka等,2016)。
慢性炎症过程涉及白细胞粘附到血管内皮并渗入受损组织。两种过程似乎都被NRF2与其编码血红素加氧酶-1(HO-1)的至少一种靶基因HMOX1一起调节。NRF2 / HO-1轴通过调节几种细胞粘附分子如血管细胞粘附分子1的表达来抑制炎性细胞与内皮的粘附(Banning和Brigelius-Flohe,2005; Wenzel等,2015)。另外,NRF2 / HO-1抑制巨噬细胞中的金属蛋白酶-9,这是组织内免疫细胞迁移所必需的(Bourdonnay等,2009)。
许多临床前研究报道,天然化合物(Satoh等,2013)或通过破坏其负调节因子KEAP1激活NRF2导致髓样白细胞(Kong等,2011)和巨噬细胞(Lin等, 2008)的强效抗炎作用。 在观察性研究中,NFE2L2中的多态性与转录活性降低相关,与炎症性肠病(Arisawa等,2008b)和慢性胃炎(Arisawa等,2007)的风险增加相关。NRF2的免疫调节作用的一个例子是在中枢神经系统。受损神经元释放出趋化因子,一种特异性激活小胶质细胞中磷脂酰肌醇3激酶/ AKT(PI3K / AKT)途径的趋化因子,导致GSK-3β的抑制和NRF2的上调(LastresBecker等,2014)。在这项研究中,来自AD和进行性核上性麻痹患者的尸检显示出趋化因子水平的补偿性增加以及上调的NRF2蛋白,表明该途径有助于限制病变大脑中的炎症反应。
B. NRF2在自身免疫性疾病中的作用
在NRF2疾病群的外围,我们发现了几种自身免疫疾病表型,如白癜风,哮喘,多发性硬化症(MS)和系统性红斑狼疮(SLE)。实际上,在实验性自身免疫性脑脊髓炎(EAE)和类风湿性关节炎(RA)的动物模型中的大量工作,以及MS和牛皮癣中的临床证据进一步指出NRF2在自身免疫疾病中的作用。氧化组织损伤和细胞凋亡可以增加自身抗原的产生,导致T细胞的活化和B细胞产生自身抗体,例如,如对3-硝基酪氨酸阳性蛋白所观察到的那样(Thomson等,2012)。此外,II相解毒酶的丢失,其中许多由NRF2转录调节,导致活性中间体的产生增加,这有助于形成半抗原或受损的大分子,有时变得具有免疫原性,从而增加引发自身免疫反应的自身抗原库。因为NRF2调节的酶在许多化学物质的解毒中起关键作用,因此可以认为NRF2可能是一种保护机制,可以抵抗环境对自身免疫发病机制的作用(Ma, 2013)。NRF2介导的自身免疫调节的潜在机制还涉及抑制促炎性Th1和Th17应答以及免疫抑制性T调节(Treg)和Th2应激的激活。还有越来越多的证据表明NRF2可以控制参与抗原呈递和适应性免疫应答调节的树突细胞(DC)和巨噬细胞的分化和功能。事实上,NRF2缺陷表明,通过增加共刺激分子的表达并因此增加抗原特异性T细胞反应性来改变DCs的功能和表型(Al-Huseini等,2013)。MS是一种慢性炎性疾病,其特征在于自身反应性免疫细胞浸润到中枢神经系统中。NRF2的缺失加剧了EAE的发展,EAE是MS的小鼠模型(Johnson等人,2010)。与NRF2缺乏相关的部分影响可能与HO-1水平降低有关。因此,具有骨髓特异性HO-1缺陷的小鼠表现出较高的病变发生率,伴随着抗原呈递细胞的活化和炎性Th17和髓鞘特异性T细胞的浸润(Tzima等人,2009)。敲除KEAP1(Kobayashi等,2016)或用各种激活NRF2的小分子治疗(Buendia等,2016)抑制了疾病的发展和严重程度。NRF2在活跃的MS病变中强烈上调,并且NRF2应答基因的表达主要在初始髓鞘破坏的区域中被发现(Licht-Mayer等,2015)。在MS脑中,NRF2及其靶标NADPH:醌氧化还原酶(NQO1)和HO-1主要在浸润性巨噬细胞中表达,在较小程度上在星形胶质细胞中表达,最有可能作为对病理性ROS形成的代偿性反应。相反,在少突胶质细胞中缺乏NRF2和抗氧化基因表达,这可能是它们在MS中的损伤和损失的基础(van Horssen等,2010)。由于免疫和氧化还原稳态的改变,MS患者外周血单核细胞中HO-1表达降低,并且在疾病恶化期间下调(Fagone等,2013)。值得注意的是,干扰素-β治疗患者的基因表达谱分析鉴定出NRF2是长期抗氧化反应和神经元保存的潜在介质(Croze等,2013)。
SLE由高氧化环境、失调的细胞死亡和去除死细胞的缺陷而突出,这导致细胞坏死作为自身抗原的来源。NRF2缺陷的雌性小鼠随年龄增长而发展为类似于SLE的多器官自身免疫性疾病,其特征为DNA氧化增加,脂质过氧化,脾细胞凋亡,抗双链DNA抗体和史密斯抗原的存在,以及伴随重要的组织损伤(血管炎,肾小球肾炎,肝炎和心肌炎)(Li等,2004)。只有雌性小鼠出现SLE进展的事实表明,雌性特异性因子可能有助于打破对自身抗原的免疫耐受(Li等,2004)。NRF2缺乏还导致CD4 + T细胞的增殖反应增强、CD4 + / CD8 +比率改变、以及SLE中促炎性Th17的促进(Ma等人,2006; Zhao等人,2016)。事实上,NRF2耗竭与狼疮性肾炎发展过程中的Th17分化和功能有关,这似乎是通过调节细胞因子信号传导抑制因子3 /磷酸化信号转导和转录激活因子(STAT)3途径和IL-1b信号来调节的(Zhao 等,2016)。此外,Nrf2-/ -小鼠的唾液腺显示出强烈的淋巴细胞浸润,使人想起Sjögren综合征,这通常与SLE有关(Ma等,2006)。SLE患者表现出氧化性DNA损伤修复机制的改变(Evans等,2000),高血清氧化蛋白水平,载脂蛋白C3(Morgan等,2007),氧化磷脂和抗氧化修饰脂蛋白的自身抗体(Frostegard等,2005)。NRF2多态性尚未与SLE易感性相关,尽管SNP rs35652124与墨西哥女性儿童期发病肾炎风险增加有关(Cordova等,2010)。
类风湿关节炎(RA)是一种全身炎症性疾病,具有复杂但仍难以捉摸的自身免疫特性,在炎症关节中,中性粒细胞、巨噬细胞和淋巴细胞被积极募集和激活。这导致促炎介质如ROS / RNS,类二十烷酸,细胞因子(IL-17,TNF-α,干扰素-?,IL-6和IL-1β)和分解代谢酶的分泌增加,这些促炎介质引发滑膜成纤维细胞的过度增殖, 关节肿胀,软骨和骨的逐渐破坏(Roberts等,2015)。NRF2基因的缺失增加了实验性RA模型中关节改变的易感性。例如,在表达T细胞受体KRN和主要组织相容性复合体II类分子A(g7) (K/BN关节炎模型)的小鼠中,以及在抗体诱导的关节炎中,NRF2缺乏加速了发病率并加重了疾病进程( Maicas等,2011; Wu等,2016b)。NRF2缺乏显着上调炎症细胞的迁移、环加氧酶-2和诱导型一氧化氮合酶的表达、ROS和RNS的产生以及促炎细胞因子和趋化因子的释放。此外,NRF2可能是关节炎中骨代谢的保护因子(Maicas等,2011),NRF2 / HO-1活化在RA动物模型和人滑膜成纤维细胞中发挥抗炎和抗氧化作用(Wu等,2016b)。有趣的是,抗风湿金(I)化合物通过激活NRF2和上调HO-1和GCLC来刺激抗氧化反应,证明其对RA的临床疗效(Kobayashi 等, 2016)。此外,NRF2 / HO-1激活介导了滑膜细胞凋亡的诱导和西洛他唑对促炎细胞因子产生的抑制(Park等,2010)以及H2S和相关化合物的抗炎作用,它们能通过对KEAP1的半胱氨酸残基进行硫化物修饰(Wu等,2016b)。其他诱导NRF2和HO-1信号传导的药物,如瑞巴派特,可以转导人和小鼠CD4+ T细胞向免疫抑制性Treg表型的分化,并通过特异性抑制STAT3来抑制TCD4+细胞向炎性Th17细胞的分化(Moon等,2014)。发炎的滑膜内过量的ROS产生似乎有助于RA的发病机制,因为患者的ROS形成,脂质过氧化,蛋白质氧化,DNA损伤和抗氧化防御机制活性降低显着增加,所有这些都促成了组织损伤和疾病进展(Datta等,2014)。响应于病理性ROS的形成,NRF2途径在RA患者的滑膜细胞和抗体诱导的关节炎小鼠的关节中被激活,但是该反应显然不足以抵消疾病进展(Wu等,2016b)。
白癜风是一种皮肤炎症性疾病,其特征在于表皮中ROS的积累,其参与黑素细胞的死亡。这些分子修饰DNA和黑素体蛋白,形成自身抗原并激活针对黑素细胞的自身免疫应答(Xie等,2016)。 遗传学研究发现NRF2启动子SNPs与白癜风易感性有关,如-650位点的SNP (Guan等, 2008),而rs35652124的C等位基因在汉族人群中被证明具有保护作用(Song等, 2016)。NRF2及其下游解毒靶基因NQO1,GCLC和GCLM在白癜风患者的表皮中上调,表明这种防御机制的激活不足(Natarajan等,2010)。
C. NRF2在慢性呼吸系统疾病中的作用
NRF2在呼吸系统疾病中的相关性在2010年得到了回顾(Cho和Kleeberger,2010),在这项工作中,我们将仅重点介绍最相关的发现(图8)。香烟烟雾是慢性阻塞性肺病(COPD)的主要危险因素。COPD患者具有功能失调的肺泡巨噬细胞,其导致不受控制的ROS产生,促炎介质,吞噬作用缺陷和一系列参与组织损伤的金属蛋白酶。慢性阻塞性肺病(COPD)患者的肺泡巨噬细胞功能失调,导致ROS生成失控、促炎介质、有缺陷的吞噬作用以及一系列参与组织损伤的金属蛋白酶。事实上,肺气肿肺组织显示实质中肺泡巨噬细胞密度与肺破坏严重程度之间存在直接关系(Finkelstein等,1995)。肺泡巨噬细胞的吞噬活性受损是引起COPD急性加重的细菌和病毒复发的主要原因,并且是发病率和死亡率的主要来源。Nrf2-/- 小鼠对香烟烟雾诱发的肺气肿表现出增强的易感性(Rangasamy等,2004)。重要的是,用异硫氰酸酯萝卜硫素(SFN)激活NRF2可恢复对细菌的识别和吞噬作用,增强肺泡巨噬细胞对肺细菌的清除,并减少野生型小鼠的炎症,但对于暴露于接触香烟烟雾的Nrf2-/-小鼠不会发生(Harvey等,2011)。在人类中,与吸烟和非吸烟的非肺气肿患者相比,吸烟相关性肺气肿患者肺泡巨噬细胞中NRF2的转录特征降低(Goven等, 2008)。NRF2表达降低与脂质过氧化产物4-羟基壬烯醛巨噬细胞表达增加有关。在NFE2L2启动子中,由三个SNPs和一个三联体重复构成的功能性单倍型,产生低至中等NRF2表达的多态性(Yamamoto等,2004),与发生COPD的风险增加有关(Hua等,2010)。低表达单倍型是发展呼吸衰竭的重要预测因子。因此,SNP rs6721961的-617A等位基因发生急性肺损伤的风险显着较高(Marzec等,2007)。病理性ROS形成可能在慢性肺纤维化的发病机制中起作用。早期研究表明,博来霉素诱导的肺纤维化在Nrf2-/- 中比在野生型小鼠中更严重(Cho等,2004)。事实上,野生型小鼠通过上调NRF2诱导抗氧化和抗炎反应,而这在Nrf2-/-小鼠中无法实现。后来证实,患有特发性肺纤维化或慢性结节病/过敏性肺炎的患者表现出NRF2的表达增加,并且支气管肺泡灌洗液中的低摩尔.wt.抗氧化剂水平增加,如尿酸,抗坏血酸,视黄醇和α-生育酚 ,表明对ROS挑战的适应性反应不成功(Markart等,2009)。机制地,NRF2缺乏增加肌成纤维细胞分化,而用SFN药理诱导NRF2导致肌成纤维细胞数量减少和转化生长因子-β(TGF-β)的促纤维化作用减弱(Artaud-Macari等人,2013)。
D. 消化系统中的NRF2
NRF2疾病组中消化系统病理表型的显着地位突出了NRF2的转录特征作为一种对异生素引发的慢性氧化损伤和炎症应激的有效适应机制的相关性。在胃肠道(GI)中,长期接触异生素可引发肠腔微生物群与免疫系统之间功能失调的相互作用(Aviello和Knaus,2017)。这可导致胃肠道的慢性疾病,如包括克罗恩病和溃疡性结肠炎的炎性肠病(IBD)表型,其中存在激活保护性NRF2应答的证据。例如,在来自IBD患者的结肠组织中,结肠上皮细胞通过与蛋白酶体蛋白表达增加相关的NRF2依赖性适应对炎症信号起反应(Kruse等,2016)。来自IBD患者的单核细胞衍生的巨噬细胞证明了特异性NRF2依赖性基因表达谱,其响应于LPS而加重,进一步表明这是一种减轻炎症反应的尝试(Baillie等, 2017)。在遗传水平上,NFE2L2基因的特定基因型(-686-684)与日本队列研究中的溃疡性结肠炎的发展相关,尤其是在女性中(Arisawa等,2008a)。事实上,GI中的病理过程高度依赖于宿主的遗传背景,与肠腔微生物群和免疫系统之间的功能失调相互作用有关(Aviello和Knaus,2017)。GI的微生物群的共生作用之一是释放适量的ROS,其引起由NRF2介导的上皮细胞和浸润的免疫细胞中的细胞保护反应(Jones等,2015)。此外,在真核生物中受NRF2转录控制的细胞保护分子也可以由共生细菌产生。例如,微生物群中的HO-1同源物可能对胃肠道内稳态有很大的贡献,这可以用于治疗一氧化碳在肠道的局部递送(Onyiah等, 2014)。
NRF2在维持胃肠道稳态方面的作用已被证实,使NRF2这一转录因子成为IBD中有希望的治疗靶点。因此,几种化合物和膳食补充剂可能表现出有益效果,如褪黑激素,3-(3-吡啶基亚甲基)-2-吲哚满酮,丁酸盐,干酪乳杆菌,左旋肉碱,4-乙烯基-2,6-二甲氧基苯酚(卡诺尔), 乳枸杞(脱脂牛奶中枸杞的配方产品)等(Orena等,2015)。因此,明确NRF2在消化道慢性和急性疾病中的参与情况,对于更好地指导NRF2通路的调节治疗方法至关重要。
肝脏也是抵抗食物异生素的第一道防线。因此,NRF2疾病组强调该转录因子在与肝损伤相关的病理表型中的相关性也就不足为奇了。早期使用Nrf2-/-小鼠模型证明其对对乙酰氨基酚诱导的肝细胞损伤、苯并[a]芘诱导的肿瘤形成以及Fas-和TNF-α介导的肝细胞凋亡具有保护作用(Aleksunes和Manautou,2007)。Nrf2-/-小鼠对化学毒性的较高敏感性与解毒酶的基础表达和诱导表达降低相关。在人类中,导致NRF2表达降低的三种NRF2启动子SNPs的功能性单倍型与胃粘膜炎症的发展显著相关,无论是独立地还是通过与幽门螺杆菌感染相互作用(Arisawa等,2007)。对原发性胆汁性胆管炎患者NRF2转录特征的分析表明,这些患者表现出NRF2表达降低以及HO-1和GCLC蛋白水平低,这些损伤在肝硬化患者中更为严重(Wasik等,2017)。
病理性ROS形成是非酒精性脂肪性肝炎(NASH)患者肝细胞损伤和疾病进展的关键机制(图8)。该疾病分两个阶段发展,一个是肝细胞中脂肪酸的逐渐积累,另一个是肝损伤和炎性病理性ROS的形成(Wang等,2018)。因此,喂食高脂肪饮食(HFD)的小鼠出现了单纯的脂肪变性,其特征是肝脏脂肪沉积增加而没有炎症或纤维化,但Nrf2-/-小鼠出现加剧的肝脏脂肪变性和大量炎症,与NASH一致(Reccia等,2017)。然而有趣的是,肝细胞特异性KEAP1的缺失在减少肝脏脂肪变性的同时,并没有改变NASH发展过程中的炎症反应,提示其具有代偿机制(Ramadori等, 2016)。至少在NASH的大鼠模型中,饮食NRF2激活剂减弱了肝纤维化的进展(Shimozono等,2013)。在NASH患者的肝脏活组织检查中病理性ROS形成的标志物增加,并且NRF2特征增加,表明其试图减少氧化剂和炎症负荷(Takahashi等人,2014)。
E. 心血管系统中的NRF2
NRF2疾病群指出心血管系统对细胞氧化还原平衡的变化以及众所周知的动脉粥样硬化、高血压和糖尿病等共病的发展具有高度敏感性(Griendling and FitzGerald, 2003a,b;Harrison等,2003;Jay等,2006)(图8)。NRF2在预防这些病理表型方面的作用已经在NRF2 -/-小鼠中得到证实,NRF2 -/-小鼠在慢性耐力运动中表现出心脏结构受损(更多的重塑事件)和功能受损(较少缩短分数)(Shanmugam等, 2017a)。它们也更容易在心肌梗塞后发生心力衰竭(Strom和Chen,2017)。相反,NRF2的组成型活化产生还原状态,其特征书是心脏GSH /谷胱甘肽二硫化物比率增加和ROS形成和丙二醛水平降低(Shanmugam等,2017b)。在人类中,Tako-Tsubo心肌病的微阵列分析表明,在这种收缩功能障碍的急性期,病理性ROS水平增加,NRF2代偿性上调(Nef等,2008)。最近,血液透析患者的全身性炎症和病理性ROS形成与NRF2的下调相关(Pedruzzi等,2015),并且两种启动子多态性(rs35652124和rs6721961)与这些患者的死亡风险增加相关(Shimoyama等,2014)。NRF2在内皮稳态中最相关的靶点之一是HO-1,其通常与铁蛋白的上调相平行,因此降低游离铁水平,并阻止芬顿型反应。由HO-1和胆绿素还原酶的联合活性产生的胆红素是清除ROS / RNS的最强大的内源性抗氧化剂之一(Jansen等,2010),并且在体外预防脂质过氧化方面非常有效( Stocker等,1987)。Hmox1-/-小鼠显示出慢性缺氧引起的肺动脉高压升高(Christou等,2000),药理学HO-1诱导可改善糖尿病并发症(Kruger等,2006),以及硝酸甘油诱导的血管功能障碍( 硝酸盐耐受性)(Wenzel等,2007)。最近,Hmox1-/-小鼠在血管紧张素II响应中显示NADPH氧化酶-2上调、血管病理ROS形成、炎症标志物、内皮功能障碍和血压升高(Wenzel et al., 2015)。事实上,高血清胆红素水平与冠状动脉疾病的发病率呈负相关(Hopkins等,1996)。胆红素阻止血管NADPH氧化酶的活化(Kwak等,1991),其参与心血管疾病的发展(Griendling和FitzGerald,2003a,b; Harrison等,2003; Jay等,2006)。外周动脉疾病是动脉粥样硬化的常见表现,患有外周动脉疾病的患者HO-1水平降低(Signorelli等, 2016)。
F. 代谢性疾病中的NRF2
2型糖尿病(T2DM)是最常见的慢性代谢疾病之一,并且在NRF2疾病组中尤为突出。胰岛素敏感组织以及胰腺中的病理性ROS形成已在T2DM患者中发现,导致胰腺β细胞分泌胰岛素和外周组织胰岛素作用严重受损(Uruno等,2015)(图 8)。同样,由于蛋白质的非酶促糖化,病理性ROS水平也有助于糖尿病并发症的发病机理。这已在糖尿病肾病中得到证实,其中肾小球表现出病理性ROS水平和NRF2的代偿性升高(Jiang等人,2010)。自2007年以来,一些研究利用动物模型和细胞系研究了NRF2在T2DM及其并发症中的作用。体外对人体细胞的研究表明,NRF2的激活是在急性高糖环境下实现的,而较长时间的培育时间或振荡的葡萄糖浓度均不能激活NRF2 (Ungvari等, 2011;Liu等, 2014)。因此,这些研究指出NRF2活化依赖于葡萄糖浓度和动力学。相反,NRF2在前驱糖尿病和糖尿病患者的外周血单核细胞中下调,表明NRF2可能是重要的治疗靶点(Jimenez-Osorio等,2014)。NRF2缺乏对高血糖的影响首先在Nrf2-/-小鼠中显示,其中氧化和亚硝化改变增强并导致早期肾损伤(Yoh等,2008)。 在随后的研究中,链脲佐菌素诱导的糖尿病Nrf2-/-小鼠表现出加剧的肾小球损伤,同时ROS产生增加和促纤维化标志物TGF-β和纤连蛋白表达的增加(Jiang等人,2010)。在这个糖尿病模型中,NRF2对血视网膜屏障功能障碍和糖尿病视网膜病变的进展具有保护作用(Xu等, 2014)。同样,HFD诱导的血管ROS水平的增加在Nrf2-/-小鼠中显着加剧,并伴有严重的内皮功能障碍,表现为乙酰胆碱诱导的主动脉松弛减弱和细胞间粘附分子-1和TNF-α表达增加( Ungvari等,2011)。
NRF2在组织特异性胰岛素抵抗中起复杂作用。因此,与野生型小鼠相比,HFD喂养的Nrf2-/-小鼠由于肝脏和骨骼肌中胰岛素信号的增强而表现出更好的胰岛素敏感性,但相反,这些小鼠由于与病理ROS形成相关的肝脂毒性过高而产生严重的NASH (Meakin等, 2014)。因此,该研究将肝胰岛素抵抗与NASH的发展分离。根据这些数据,随后的研究表明,由于CYP2A5酶的表达减弱,HFD喂养的Nrf22-/-小鼠的肝脏由于GSH的显着消耗而表现出更高的病理性ROS形成(Cui等人,2013)。NRF2在肝细胞中的敲低增强了棕榈酸诱导的细胞凋亡,棕榈酸是一种在胰岛素抵抗性肥胖患者中高度升高的脂肪酸。这种效应与病理性ROS的产生增加相关,再次强化了NRF2在NASH进展中的关键作用(Pilar Valdecantos等,2015)。
为了进一步研究NRF2在代谢综合征中的作用,我们在瘦素缺乏(ob/ob)小鼠模型中去除NRF2,该模型具有极好的正能量平衡(Xue等, 2013)。有趣的是,全身ob / ob / Nrf2-/-小鼠或脂肪细胞特异性ob/ob/ Nrf2-/-小鼠显示白色脂肪量减少,显示NRF2是脂肪生成的关键参与者。这些小鼠具有更严重的代谢综合征,其特征在于高脂血症,加重的胰岛素抵抗和高血糖,表明代谢综合征与病理性ROS形成之间的机制联系。
另一组研究评估了持续诱导NRF2在葡萄糖代谢中的作用。NRF2基因诱导利用Keap1的一个亚型等位基因(Keap1flox/- 突变体)降低肥胖糖尿病db/db小鼠的血糖,是通过抑制肝细胞中cAMP-CREB信号传导以及其他糖异生基因(例如过氧化物酶体增殖激活受体辅激活因子-1α)来抑制肝葡萄糖6磷酸酶(Uruno等,2013)。此外,Keap11敲低小鼠中NRF2活性的增强增加了肝脏中AMP活化蛋白激酶(AMPK)的磷酸化,以及骨骼肌中的胰岛素信号传导,导致葡萄糖耐量的显着改善(Xu等,2013)。由于NRF2在T2DM背景下的多效活性,所有这些和其他研究的结果证明需要设计多种遗传和药理学策略来阐明参与全身葡萄糖稳态控制的组织中的全部NRF2功能。
除了年龄,体重和血糖等糖尿病因素外,与NRF2相关的遗传因素在人类中研究较少。在中国人群中,SNP rs6721961与病理性ROS形成和新诊断的T2DM风险有关,并且还可能导致胰岛素分泌能力受损和胰岛素抵抗增加(Wang等,2015)。在墨西哥混血男性中,相同的SNP与糖尿病相关(Jimenez-Osorio等,2017)。在一项汉族志愿者参与的病例对照研究中,发现T2DM患者无论有无并发症,包括周围神经病变、肾病、视网膜病变、足溃疡和微血管病变,NFE2L2基因4个SNPs的基因型和等位频率存在显著差异(Xu等, 2016b)。
G. 神经退行性疾病中的NRF2
NRF2疾病组提供NRF2参与几种神经退行性疾病的证据,包括AD和PD,其代表老年人最普遍的认知和运动障碍。在神经退行性疾病中,低级病理性ROS形成与蛋白质稳态之间的联系尤为重要,因为大多数这些病理表型的特征在于特定蛋白质的异常聚集(图8)。在细胞和动物模型中提供了指向蛋白病变中病理ROS形成以及NRF2作为蛋白酶体和自噬调节因子的证据(Pajares等, 2017)。最初,据报道自噬货物蛋白质sequestosome 1(SQSTM1)与NRF2竞争结合KEAP1。最初,有报道称自噬货物蛋白sequestosome 1 (SQSTM1)与NRF2竞争与KEAP1的结合。SQSTM1将KEAP1置于自噬体降解途径,因此上调NRF2(Komatsu等,2010)。SQSTM1将KEAP1置于自噬体降解途径,因此上调NRF2(Komatsu等,2010)。最近,发现NRF2调节参与自噬起始、货物识别、延伸和自溶酶体清除的自噬基因的表达(Pajares等,2016)。在本研究中,Nrf2-/-小鼠中,转基因过表达人突变淀粉样前体蛋白和tau蛋白导致淀粉样病变和tau蛋白病加重。NRF2缺乏症和神经退行性变之间的联系被越来越多的动物模型证据所支持(Johnson and Johnson, 2015)。一般的观点是受损的神经元试图激活NRF2依赖性转录,可能是为了增加它们自身的存活率。此外,星形胶质细胞中NRF2的上调参与代谢补偿,包括增加GSH的供应以增强其增殖能力(Bolanos,2016),而小胶质细胞中的NRF2上调使该免疫细胞恢复到静止状态(Rojo等,2014a)。Ramsey等人(2007)证明了NRF2在PD患者的多巴胺能神经元中的核定位。其他研究发现淀粉样蛋白前体蛋白和tau损伤神经元表达NRF2及其靶SQSTM1水平升高,可能是通过自噬清除这些毒性蛋白的补偿机制(Lastres-Becker等,2014; Pajares等,2016)。与这些结果一致,AD和PD脑中HO-1、NQO1、GCLM和SQSTM1水平升高(van Muiswinkel等, 2004;Cuadrado等,2009;Schipper等,2009;LastresBecker等,2016)。此外,与NRF2表达相关的细胞保护蛋白,如NQO1和SQSTM1,在路易体中被部分隔离,表明PD患者中NRF2特征的神经保护能力受损(Lastres-Becker等,2016)。这种差异的一种可能解释是NRF2及其靶基因水平可能在衰老和疾病进展过程中发生变化。
NFE2L2的一些SNP单倍型与ALS,AD或PD的风险降低或延迟发作相关。在关于先前与高基因表达相关的三种功能性启动子SNPs的两项研究中分析了ALS的发病。有趣的是,这种单倍型与ALS发病延迟4年有关(Bergstrom等,2014),但另一项研究未发现明确的相关性(LoGerfo等,2014)。关于AD,一个单倍型等位基因与AD的2年早期发病相关,表明NFE2L2基因的变体可能影响AD进展(von Otter等,2010b)。已经更详细地分析了NFE2L2与PD的遗传关联。NFE2L2启动子中的三个SNPs(rs6721961,rs6706649和rs35652124)在病例对照研究中被证明为保护性单倍型(von Otter等,2010a)。这种单倍型在瑞典队列中延迟了疾病的发作,甚至降低了波兰队列中PD的风险。这些结果得到了四项新的独立的欧洲病例对照研究的支持(von Otter等,2014),但未在台湾人群中复制(Chen等,2013),这表明种族和环境因素存在差异。另一种方法是将来自嗅黏膜的PD细胞暴露于烟提物或杀虫剂中,以评估基因-环境的相互作用,并且鉴定了几种影响对这些毒素的易感性的SNPs(Todorovic等,2015)。总而言之,NRF2的轻微激活(例如对于NFE2L2基因的某些功能性单倍型所发现的)应该足以触发大脑中的保护机制。
IV. 癌症中Keap1的悖论
NRF2在肿瘤发生和进一步肿瘤进展中的作用似乎存在明显的二分法。一方面,通过激活生物转化反应,NRF2可以防止化学诱导的癌症发生。临床前研究表明NRF2在大鼠体内经药理激活后,对黄曲霉毒素B(1)诱导的肝癌具有完全的保护作用(Johnson等, 2014)。相反,NRF2引发的保护性反应在确定的癌症中提供了生长优势,这将是本节的重点。不断增加的ROS水平可以通过改变基因组稳定性来维持肿瘤发生,同时激活特定的氧化还原信号转导和促进肿瘤细胞存活和增殖的炎症过程(Reuter等,2010)。因此,NRF2的上调代表癌细胞适应的机制,以耐受推动肿瘤进展的高ROS水平(Schumacker,2006)以及维持导致肿瘤复发和远处转移形成的癌症干细胞(Ryoo等,2016)。例如,来自人类结肠直肠肿瘤的癌症干细胞中的NRF2特征指出了由高水平的GCLC,谷胱甘肽过氧化物酶和硫氧还蛋白还原酶-1介导的保护机制,这是这些细胞抵抗应激物和化学治疗剂的能力的基础(Emmink等,2013)。从这个角度来看,NRF2在癌细胞中表现得像一个致癌基因,通过诱导ARE介导的细胞保护反应的慢性激活,可以帮助其适应其氧化环境(Panieri和Santoro,2016)。已经报道了NRF2的恶性激活的几种机制,包括体细胞突变,表观遗传学和致癌信号传导改变。
据报道,沿着NFE2L2的编码序列在癌症中接近600个体细胞突变(Gao等人,2017)。在图9中,我们显示了来自10,000名癌症患者的数据集的结果(Zehir等人,2017)。在大多数情况下,这些突变改变了NRF2的DLG和ETGE基序与KEAP1的相互作用,从而诱导NRF2在几种实体瘤(包括食道癌,皮肤癌,肺癌和喉癌)中的过度活化(Kim等,2010b; Taguchi和 Yamamoto,2017)。例如,在晚期食管鳞状细胞癌中,NRF2功能获得性突变与肿瘤复发和预后不良相关,其原因包括增殖增加、独立依附生存以及对化疗和放疗的抵抗(Shibata 等, 2011)。KEAP1基因中的功能丧失突变在一些实体肿瘤例如肺癌中也是常见的(Singh等人,2006)。基于这一通路调控β-TrCP/NRF2的有力证据,奇怪的是在NRF2与β-TrCP的交界面未发现干扰体突变。这一事实表明,由于未知的原因,这些突变是不可生存的,或者β-TrCP逃逸导致NRF2水平的升高不足以驱动致癌性。
尽管如此,体细胞突变仅在一小部分癌症患者中引起长期/慢性NRF2激活。在基因表达水平上,有趣的是位于人NRF2基因的ARE增强子的SNP rs6721961(-617C>A)的等位基因消除了NRF2的自身诱导,这与这些癌症患者的显着存活相关(Okano等,2013)。已经在肺肿瘤中描述了由于KEAP1的三个CpG位点的启动子高甲基化引起的表观遗传变化,导致随后的NRF2活化,其可以通过5-氮杂-2’-脱氧胞苷处理逆转NRF2的活化(Wang等,2008)。最近已经回顾了miRNAs在NRF2水平的转录后调节中的作用(Kurinna和Werner,2015)。简而言之,miR200a靶向人乳腺癌细胞中的KEAP1 mRNA,导致其降解并随后激活NRF2(Eades等,2011)。反过来,miR28促进NRF2 mRNA的降解(Yang等人,2011)。
致癌基因或突变的肿瘤抑制因子可以增强癌症中NRF2的活化。KRAS,BRAF或c-MYC的内源性致癌等位基因上调NRF2,可能是通过致癌基因介导的ROS和随后的KEAP1慢性失活(DeNicola等,2011)。肿瘤抑制因子p53的突变形式,通过增强营养摄取和构建块的合成来维持癌细胞的生长,也可以通过与结合NRF2启动子的Sp1转录因子的串扰来上调NRF2(Tung等,2015)。
各种蛋白激酶对NRF2(磷酸-NRF2)的磷酸化是107种肝细胞癌中潜在的激活机制。增加的磷酸化NRF2水平与表现出这种独特表型的患者的KEAP1表达降低和5年总体存活率差有关(Chen等,2016)。此外,磷酸酶和张力蛋白同源物(PTEN)肿瘤抑制因子的突变维持过度活跃的和致癌的磷脂酰肌醇-3-激酶(PI3K)-AKT信号传导,并由于为NRF2蛋白酶体降解的PTEN / GSK-3 / β-TrCP途径的下调而导致NRF2活性增加。(Rada等,2011,2012; Cuadrado,2015)。针对PTEN / GSK-3 / β-TrCP途径的治疗干预措施应考虑到GSK-3既可作为肿瘤抑制因子又可作为肿瘤启动子,并且还与癌症干细胞的产生有关(McCubrey等,2014)。
一些应激诱导的蛋白质与KEAP1相互作用,因此在癌细胞中,这些蛋白与KEAP1结合的NRF2竞争。因此,NRF2逃脱了KEAP1介导的降解。NRF2与KEAP1结合的竞争对手之一是自噬衔接蛋白SQSTM1的磷酸化形式,该蛋白发生在癌细胞用于维持自身生长的选择性自噬过程中(Shimizu等, 2016)。还证明了促进癌症干细胞中细胞周期停滞的细胞周期蛋白依赖性激酶抑制剂p21通过其KRR基序与NRF2中的DLG和ETGE基序的相互作用抑制NRF2与KEAP1的结合(Chen等人,2009)。最近,显示具有ETGE基序的二肽基肽酶3可以与NRF2竞争结合KEAP1(Hast等人,2013)。可能由氧化还原状态的长期改变诱导的二肽基肽酶3的过表达与ARE基因的表达增加和预后不良相关,特别是在雌激素受体阳性乳腺癌中(Lu等人,2017)。
NRF2诱导导致癌症进展的代谢变化。例如,一项多平台非靶向代谢组学研究确定了乳腺肿瘤样本中代谢物变化的模式(Tang等,2014)。他们发现GSH和3-(4-羟基苯基)乳酸通过与NRF2的相互作用与BRCA1参与氧化还原稳态呈正相关。代谢组学研究还表明,NRF2可以增加癌细胞中的有氧糖酵解,以支持其高能量需求。这通过NRF2介导的锰-超氧化物歧化酶表达的诱导发生,导致线粒体过氧化氢的产生升高和AMPK的活化。该过程由caveolin-1调节,其直接结合NRF2和KEAP1,并阻碍NRF2活化并因此阻碍糖酵解转变。这显然解释了为什么通常更具侵略性的糖酵解肿瘤具有caveolin-1低/ Mn超氧化物歧化酶高的表型(Hart等,2016)。NRF2还可以驱动葡萄糖和谷氨酰胺向肿瘤细胞增殖所需的合成代谢途径(Mitsuishi等,2012)。在PI3K-AKT信号通路活跃和KEAP1活性缺失的情况下,NRF2被证明可以诱导癌症中葡萄糖代谢从糖酵解向合成代谢途径(嘌呤合成)的转移(Mitsuishi等, 2012;Xu等, 2016a)。以上这一点被强调通过NRF2介导的参与磷酸戊糖途径的基因转录和NADPH(葡萄糖6磷酸脱氢酶;磷酸葡萄糖酸脱氢酶,苹果酸酶1,异柠檬酸脱氢酶1,转酮醇酶和转醛酶)的产生以及涉及嘌呤核苷酸合成(磷酸核糖焦磷酸酰胺转移酶,亚甲基四氢叶酸脱氢酶2)的相关基因。
NRF2的激活赋予癌细胞生长优势的事实可能会争辩说,在这项工作中提到的慢性疾病NRF2的药理学活化可能意味着患癌症的高风险。然而,必须考虑到NRF2的致癌活性需要其基因或KEAP1发生突变,从而导致NRF2信号的持续高水平诱导。但在药理学治疗中并非如此,在药理学治疗中,调节药物剂量和NRF2活性是可能的。此外,经验证据表明,参与NRF2激活剂临床试验的受试者没有显示出更高的癌症风险。自从2013年获得监管机构批准以来,已经服用NRF2活化剂富马酸二甲酯数年的MS患者就是最好的例证。相反,在癌症患者中使用NRF2抑制剂可能导致NRF2疾病组中描述的病理表型的表现。当NRF2抑制剂到达临床时,这种可能性需要进一步研究。
V. NRF2药物组
本节试图开发一种NRF2药物组,该药物组可用于未来的临床指导,以NRF2疾病组的病理表型为中心靶向NRF2进行治疗。如图2B中所述,通过靶向KEAP1来正在寻求NRF2的药理学活化以增加其稳定性。这些策略基于发现改变KEAP1结构的亲电化合物或阻止NRF2与KEAP1对接的小分子。虽然尚未经验证实,但GSK-3抑制剂应阻止b-TrCP识别NRF2,并且可发现一些化合物以防止NRF2与b-TrCP的结合。用靶向bZip二聚化结构域的化合物分析NRF2的抑制作用,以防止形成活性NRF2 / MAF异二聚体。通过与其他转录因子比较,可能发现阻碍NRF2 / MAF异二聚体与ARE结合的小分子(图9C)。本节总结了新药物发现和重新调整药物用途的转化观点中最重要的发现。 A. 亲电NRF2诱导剂
大多数已知的生理学或药理学NRF2诱导剂是亲电分子,其通过氧化或烷基化共价修饰富含硫醇的KEAP1蛋白中存在的半胱氨酸残基(Hur和Gray,2011; Satoh等,2013)。KEAP1是最适合作为亲电子或氧化还原传感器的蛋白质之一,因为它在人体中含有27个半胱氨酸残基,并起到亲电子陷阱的作用。KEAP1的半胱氨酸C151,C273和C288似乎是最容易发生亲电子反应的(图2B),尽管存在一些特异性(Yamamoto等,2008; Saito等,2015)。亲电子加合物以两种不同的方式抑制KEAP1。 一种是诱导KEAP1的构象变化,这将导致其对NRF2的结合能力丧失。另一个是阻断KEAP1和CUL3 / RBX1之间的相互作用,导致KEAP1与NRF2的隔离以及新合成的NRF2的进一步稳定化(Rachakonda等,2008; Baird和Dinkova-Kostova,2013; Cleasby等, 2014; Saito等,2015)。NRF2调制剂的至少30项近期专利在世界国际产权组织中被收录。这些专利正在保护查尔酮衍生物,新型酰胺三萜衍生物,氘取代的富马酸衍生物,3-烷基氨基-1H-吲哚丙烯酸酯衍生物,甲醇酰胺,含有活化乙烯基的苄基衍生物,穿心莲内酯或[S]+阿扑吗啡,以及倍半萜内酯衍生物(Sun等,2017)。尽管从临床前概念验证的角度来看,大多数这些化合物在某种程度上被证明是有用的,但它们的临床价值迄今通常非常有限。其中只有少数药物进入了临床试验,而食品和药物管理局(Food and Drug Administration)或欧洲药品管理局(European Medicines Agency)等监管机构批准的药物更少。我们在这项研究中讨论了转化管道中最发达的NRF2激活剂。
富马酸酯是KEAP1改性剂的最突出的例子,并且富马酸二甲酯(DMF)是迄今为止唯一的食品和药物管理局和欧洲药品管理局批准的药物,其注册为NRF2活化剂。单酯形式的DMF,富马酸单甲酯(MMF)被描述为其活性代谢物。DMF和MMF是迈克尔受体,其直接与KEAP1中存在的半胱氨酸残基反应(Lin等,2011)。
从NRF2的功能尚不清楚开始,DMF和其他富马酸酯已经用于治疗牛皮癣超过50年。该化合物在欧洲以商品名Fumaderm获得许可。临床试验显示,在DMF治疗12-16周后,银屑病面积和严重程度指数降低至50%-80%(Altmeyer等,1994; Mrowietz等,1998)。最近,在III期试验(BRIDGE)中,DMF已证明其在治疗患有中度至重度慢性斑块状银屑病的成人中的功效(Mrowietz等,2017)。富马酸盐在银屑病病变缓解中的作用机制包括外周T细胞数量的减少以及从Th1向Th2免疫应答的转变(Ghoreschi等,2011; Tahvili等,2015)。在另一种自身免疫性疾病SLE中,富马酸酯已被用作治疗严重、广泛和难治性皮肤表现的全身联合疗法(Saracino和Orteu,2017)。
2013年,DMF以商品名Tecfidera被批准用于治疗MS(Xu等,2015)。在MS患者中使用DMF是通过在EAE的MS小鼠模型中获得的阳性结果推动的。对疾病进程和组织学的显着治疗效果与脊髓中巨噬细胞介导的炎症显着减少有关。血液中的多重细胞因子分析证明DMF处理的动物中抗炎细胞因子IL-10的增加(Schilling等人,2006)。此外,DMF还改善了野生型中髓鞘,轴突和神经元的保存,但在Nrf2-/-小鼠中没有(Ellrichmann等,2011)。在人类中,DMF显示MS病变和年复发率显着降低(Schimrigk等,2006)。两项III期临床试验DEFINE和CONFIRM证实了这些结果(Fox等,2012; Gold等,2012)。因此,DMF目前被用作复发缓解型MS的第一线治疗,其不能通过传统疗法治疗。新的DMF配方正在测试和申请专利,以提高药物的生物利用度和功效(Sun等,2017)。例如,MMF已被用于开发第二代NRF2诱导物作为前药(Zeidan等,2014)。铅化合物ALKS-8700,一种MMF的2-(2,5-二氧代-1-吡咯烷基)乙酯衍生物,在体内迅速转化为MMF,因此提高了其生物利用度并减少了胃肠道副作用。ALKS-8700目前正在进行III期临床试验(EVOLVE MS)。
DMF和MMF调节免疫应答。例如,它们通过减少炎性细胞因子的释放并因此减少DCs处理抗原的能力来抑制DCs的成熟。此外,DMF和MMF激活天然杀伤细胞以裂解DCs并增强DCs和T细胞的凋亡(Ghoreschi等,2011; Al-Jaderi和Maghazachi,2015)。因此,DMF和MMF阻碍T细胞介导的自身反应。一些研究表明,DMF还通过触发GSH耗竭诱导II型DCs,这导致HO-1活性增强和STAT1磷酸化抑制。这些经典的II型DCs抑制Th1和Th17介导的反应,支持Th2的反应。此外,通过DCs增加IL-10的产生有利于CD4+T细胞向抑制性Treg表型的分化(Ockenfels等,1998; Ghoreschi等,2011)。DMF还抑制NF-κB的核转位(Peng等,2012),从而在小胶质细胞和星形胶质细胞(Brennan等,2017)以及外周血单核细胞(Eminel等,2017)中产生炎症介质,如TNF-α,IL-1β,IL-6,趋化因子,粘附分子和一氧化氮。此外,DMF发挥抗血管生成作用,其依赖于内皮细胞中血管内皮生长因子受体-2表达的下调(Meissner等,2011)。最近的研究结果表明,DMF减少了CD4+,CD8+,Th1和Th17细胞的数量,而CD4+/CD8+比例和Th2亚群在这些患者的血液中增加。有趣的是,DMF/MMF对T细胞活化的抑制作用主要局限于记忆T细胞(Wu等, 2017)。DMF或MMF的这些免疫调节活性对于保护少突胶质细胞免受ROS诱导的细胞毒性具有重要意义(Scannevin等,2012)。
其他机制可以解释NF-κB的抑制,而与NRF2活化无关。因此,DMF可能与几种调节NF-kB信号传导的蛋白质中的半胱氨酸残基相互作用(Blewett等,2016)。此外,DMF可以抑制泛素偶联酶,从而防止NF-κB的IkB抑制因子在IL-1β或Toll样受体激动剂的作用下降解(McGuire等, 2016)。此外,DMF直接结合蛋白激酶C-θ中的特定半胱氨酸残基,这是参与T细胞受体信号传导的关键激酶(Blewett等,2016)。此外,MMF和DMF激活羟基羧酸受体-2,导致NF-κB的抑制和促炎细胞因子和粘附分子的下调(Chen等,2014; Gillard等,2015)并导致中性粒细胞浸润减少 (Chen等,2014)。尽管这些NRF2非依赖性作用与EAE的急性炎症期相关,但DMF在慢性自身免疫性脱髓鞘中的神经保护功效取决于NRF2活化(Linker等,2011)。在Nrf2-/-和野生型小鼠中DMF治疗的临床益处与炎性Th1和Th17细胞的减少以及抗炎M2单核细胞的诱导相关。同时,在野生型中观察到CD80和CD86共刺激分子的表达降低,但在Nrf2-/-小鼠中未观察到,表明至少这些作用是NRF2依赖性的(Schulze-Topphoff等,2016)。
DMF用于治疗自身免疫疾病的成功表明,具有由慢性,低级炎症和病理性ROS形成强调的共同病理机制的其他疾病可能受益于该药物的重新定位。在亨廷顿舞蹈病的小鼠模型中,DMF治疗保留了存活率、肌肉功能和体重,这与完整神经元数量的增加有关(Ellrichmann等,2011)。此外,在最近的PD临床前研究中,使用该疾病的α-突触核蛋白病模型,由于自噬诱导受损,DMF在野生型中具有神经保护作用,但在Nrf2-/-小鼠中则不具有神经保护作用(Lastres-Becker等,2016)。
DMF被证明可预防糖尿病小鼠的内皮功能障碍和心血管病理性ROS形成和炎症(Sharma等,2017),并且在注射链脲佐菌素后载脂蛋白E缺陷小鼠中动脉粥样硬化、肾功能不全和其他糖尿病并发症减少(Tan等,2014)。此外,一些研究表明,DMF可能通过抑制NF-κB途径发挥抗肿瘤活性,因此在治疗侵袭性癌症中增加了治疗价值(Kastrati等,2016)。DMF是网络药理学方法中老药新用概念的相关实例。
合成三萜类化合物是2-氰基-3,12-二氧代 - 油酸-1,9(11) - 二烯-28-酸盐(CDDO; 巴多索隆,RTA401)的衍生物,其类似于天然产物齐墩果酸。它们通过其α-β不饱和支架表现出迈克尔受体活性,并且代表了最有效的NRF2诱导剂(Sun等,2017)。它们与KEAP1的C151相互作用并阻碍其与CUL3的相互作用,从而导致NRF2激活(Cleasby等,2014)。原理证明研究强烈支持合成的三萜类化合物用于退行性疾病,并且正在成为Reata / Abbott作为炎症抗氧化剂调节剂的深入研究的焦点。例如,CDDO-咪唑(CDDO-Im RTA403)在野生型而非Nrf2-/-小鼠的腹膜中性粒细胞中诱导各种抗氧化基因(Hmox1,Gclc,Gclm和Nqo1)的表达,并减弱LPS诱导的ROS产生和产生和促炎细胞因子的产生,从而降低死亡率(Thimmulappa等,2006b)。CDDO-乙基酰胺(RTA405)和CDDO-CDDO-三氟乙基酰胺(RTA 404)在毒素诱导的PD模型(1-甲基-4-苯基-1,2,3,6-四氢吡啶)中测量的所有终点均具有显着作用 (Kaidery等,2013)。在MS的EAE模型中,CDDOCDDO-三氟乙基酰胺抑制炎症、病理性ROS形成和髓鞘变性(Pareek等,2011)。
CDDO-甲酯(CDDO-Me,RTA 402)是在用于治疗糖尿病肾病的临床试验中达到的第一个CDDO(Pergola等,2011)。虽然第二阶段的结果非常令人鼓舞,但由于心血管安全问题(Zhang,2013),CDDO-Me后来在第三阶段(BEACON试验)被撤回,这与NRF2无关,但最有可能是脱靶改变内皮素信号传导(de Zeeuw等,2013; Chin等,2014)。目前,CDDO-Me正在进行临床研究,作为Alport综合征和肺动脉高压的潜在治疗方法(表2)。为了改善其安全性,进一步的研究导致了CDDO-二氟丙酰胺(RTA-408, omaveloxone)的开发,该药物目前正在进行二期试验,用于治疗弗里德雷希的共济失调、眼部炎症和眼部手术后疼痛。
奥替普拉是一种有机硫化合物,用作抗血吸虫病药物,目前正在进行III期试验,用于治疗非酒精性脂肪性肝炎。用于治疗亨廷顿氏病的高级临床试验正在开发中,米诺环素是一种抗生素,由于NRF2活化而具有神经保护作用(Kuang等,2009)。用于治疗急性肾病的I期临床研究中的另一种NRF2诱导剂是CXA-10,一种通过激活NRF2具有抗炎特性的硝基脂肪酸(Batthyany和Lopez,2015)。近年来,许多具有相同作用机制的NRF2诱导剂已被描述(Buendia等, 2015a,b, 2016),其中一些还处于临床前研究阶段,如化合物VEDA-1209,它是一种查尔酮衍生物,具有良好的抗炎作用,可用于溃疡性结肠炎的治疗。
SFN是由有机硫化合物萝卜硫苷的酶促裂解产生的异硫氰酸酯,其存在于西兰花、卷心菜和其他十字花科植物的芽苗中。催化反应由植物中发现的黑芥子酶和胃肠道的微生物群驱动(Kensler等,2013)。催化反应由植物中发现的黑芥子酶和胃肠道的微生物群驱动(Kensler等,2013)。最近,SFN已经通过化学合成获得(Kim等,2015)。通过向患有T2DM的患者施用含有SFN的花椰菜芽粉,实现了SFN向临床的转化(Bahadoran等,2012)。西兰花粉降低血浆丙二醛和氧化低密度脂蛋白(LDL),提高总抗氧化能力。心血管危险因素如血清甘油三酯,氧化LDL/LDL比率和血浆致动脉粥样硬化指数(甘油三酯对数/高密度脂蛋白比率)也降低。此外,促炎标志物如C-反应蛋白和IL-6减少。在最近的一项研究中,SFN作为浓缩西兰花芽提取物使用,通过NRF2的核转位抑制肝细胞葡萄糖的生成,并降低参与糖异生的关键酶的表达。此外,SFN降低了患有T2DM的肥胖患者的空腹血糖和糖化血红蛋白(Axelsson等,2017)。SFN诱导的NRF2活化通过降低ROS负荷和抑制NF-κB和TGF-β1信号传导途径来保护肾细胞免于狼疮性肾炎(Jiang等,2014a)。
关于神经退行性疾病,已经表明SFN穿过血脑屏障并提供足够的脑生物利用度来激活NRF2特征并减少LPS诱发的神经炎症,这反映在促炎性标志物(诱导型一氧化氮合酶、IL-6、TNF-α)的减少中和海马中的小神经胶质细胞增生(Innamorato等,2008)。SFN还保护多巴胺能神经元免受帕金森病毒1-甲基-4-苯基-1,2,3,6-四氢吡啶的影响,并减弱星形胶质细胞增生和小神经胶质细胞增生(Jazwa等,2011)。与这些发现一致,SFN降低了磷酸化tau水平并增加了Beclin-1和LC3-II,表明NRF2活化可能通过大脑中的自噬促进这种毒性蛋白的降解(Jo等,2014)。SFN处理的脊髓损伤大鼠炎症因子水平明显降低,挫伤体积减小,协调性改善(Wang等, 2012)。该药还通过保留血脑屏障和减少病理性ROS形成和炎症细胞数量来改善EAE(Li等,2013)。SFN迄今已用于至少32项针对慢性疾病的临床研究,如癌症、哮喘、慢性肾病、T2DM、囊性纤维化、自闭症和精神分裂症(Duran等,2016; Houghton等,2016)(表2)。
表2. 选择的NRF2诱导剂作为KEAP1的亲电子修饰剂
该引用对应于ClinicalTrials.gov中的代码。
总之,这些观察结果为其他SFN衍生化合物的开发铺平了道路,这些化合物表现出更好的药代动力学特征。SFN是一种在亲水性介质中稳定性较差的油性物质。其物理化学特征促使Evgen Pharma(Wilmslow,Cheshire,England)开发出一种环糊精复合物制剂Sulforadex,该制剂正在进行用于治疗蛛网膜下腔出血的II期临床试验。SFN还与褪黑激素杂交以产生ITH12674,该化合物被设计为具有用于治疗脑缺血的双重药物-前药作用机制(Egea等人,2015)。
姜黄素是姜黄中发现的主要类姜黄素,已被用于治疗肥胖症、代谢综合征和前驱糖尿病。采用气相色谱-电子冲击质谱法对姜黄素对大鼠肝脏的影响进行了非靶向代谢组学研究。间断摄入姜黄素可上调NRF2,并在抗肝损伤方面发挥抗氧化和抗炎作用(Qiu等,2016)。口服姜黄素可有效降低血清甘油三酯、IL-1β、IL-4和血管内皮生长因子,以及增加血液中脂联素水平。在T2DM患者中,姜黄素降低空腹血糖,糖化血红蛋白,血清游离脂肪酸,甘油三酯和尿酸的水平,并增加脂蛋白脂肪酶的水平(Na等,2013; Chuengsamarn等,2014)。
白藜芦醇是一种保护植物免受真菌感染的多酚,存在于葡萄皮、红葡萄酒、浆果和许多其他植物中。白藜芦醇通过下调KEAP1表达和激活蛋白去乙酰化酶sirtuin-1来激活NRF2信号通路,从而发挥抗氧化作用(Ungvari等, 2010)。在健康受试者中,白藜芦醇的饮食施用防止了血浆中胆固醇、内毒素、促氧化剂和炎性标记物(p47phox,KEAP1,IL-1β和TNF-α)的升高。这些事件与NRF2活性的升高相关,NRF2活性升高是由NRF2靶标NQO1和谷胱甘肽S-转移酶的表达增强所确定的(Ghanim等,2011)。在T2DM患者中,治疗4周后胰岛素敏感性得到改善,这通过AKT增强胰岛素信号传导,病理性ROS形成减少和糖化血红蛋白水平降低来确定(Brasnyo等,2011; Bhatt等,2012)。总的来说,白藜芦醇在动物模型和患者中可预防高血压、高胆固醇血症、动脉粥样硬化、缺血性心脏病、糖尿病和代谢综合征等主要心血管、炎症、氧化和代谢并发症(Xia等, 2017)。
经常被忽视的问题是亲电KEAP1抑制剂缺乏选择性。亲电试剂与细胞中不同的亲核试剂发生反应,从而表现出非靶向和非预期的副作用。例如,CDDO-Im可与500多种不同的目标相互作用(Yore等,2011)。通常,几种蛋白磷酸酶在其催化中心含有对氧化还原敏感的半胱氨酸,并且一些KEAP1抑制剂可以修饰和灭活这些磷酸酶,因此扰乱信号传导网络。这些磷酸酶之一是PTEN(Lee等,2002; Kitagishi和Matsuda,2013; Han等,2015)。PTEN的催化C124残基可以通过与强亲电试剂例如CDDO-Im(Pitha-Rowe等,2009)和叔丁基氢醌(Rojo等,2014b)形成加合物来修饰。然后,PI3K/AKT通路的激活增加涉及GSK-3的抑制和NRF2随后的稳定(图2C) (Rada等, 2011, 2012)。此外,KEAP1与其他同样含有高亲和力结合基序ETGE的蛋白相互作用(Hast等, 2013),如Bcl-2和IKKβ (Kim等, 2010a;Cazanave等,2014)。因此,从KEAP1缺陷细胞获得的一些结果可能不一定与NRF2活化有关。
B. 用于NRF2激活的蛋白质-蛋白质相互作用抑制剂
为了克服选择性的缺陷,出现了一类阻止NRF2与KEAP1对接的新型NRF2诱导剂(Richardson等,2015)。通过先前阐明KEAP1(Padmanabhan等人,2006)的X射线晶体结构与含有NRF2的高亲和力结合ETGE基序的肽结合,已经实现了PPI抑制剂的使用(Lo等,2006)。KEAP1包含六叶β螺旋桨,具有特定的疏水和亲水残基,参与采用β-发夹结构的EGGE基序的对接。对接主要受KEAP1的几个精氨酸与ETGE基序中的两个谷氨酸之间的静电相互作用的支持(Lo等人,2006; Padmanabhan等人,2006)。NRF2的低亲和力DLG基序与KEAP1的对接也被表征(Tong等, 2007)。基于这些相互作用,拟肽化合物是PPI抑制剂的第一个例子,其对亲电试剂的选择性显着提高(Hu等,2013; Marcotte等,2013; Winkel等,2015)。这些抑制剂在细胞中表现出较弱的活性,现在已经报道了一种新的基于循环肽的激发策略。这些肽之一显示出对KEAP1的高结合亲和力和NRF2的激活,并在小鼠巨噬细胞中引发抗炎作用(Lu等,2018)。最近对KEAP1/NRF2相互作用的新多肽和小分子抑制剂的发现进行了综述(Abed等, 2015;Jiang等, 2016)。简而言之,最初使用表面等离振子共振和荧光偏振测定评估一系列截短的NRF2肽作为PPI的直接抑制剂(Hu等,2013)。抑制能力最小的肽序列为LDE-ETGE-FL的9-mer序列(Chen等, 2011;Inoyama等,2012)。与此同时,Wells和合作者(Hancock等,2013)使用噬菌体展示文库结合高通量荧光偏振测定法搜索新推定的肽配体。他们发现,与单独的天然肽相比,基于NRF2和SQSTM1的ETGE基序的杂合肽对KEAP1具有更高的结合活性。为了促进细胞摄取,设计一种肽,其中ETGE基序与HIV-Tat蛋白的细胞转导结构域和钙蛋白酶的切割序列(DEETGE-Cal-Tat)融合。该肽在脑缺血的小鼠模型中显示出神经保护和认知保护作用(Tu等,2015)。
已经描述了五个PPI抑制剂家族:四氢异喹啉(Jnoff等,2014; Richardson等,2015),硫嘧啶(Marcotte等,2013),萘(Jiang等,2014b),咔唑酮(Ranjan等,2014)和尿素衍生物(Sato等,2013)。表3汇总了最近针对这些小分子的专利。 尽管这些化合物非常有前途,但仍需要证明它们对KEAP1/NRF2相互作用具有选择性,因为KEAP1也至少靶向Bcl2和IKK(Kim等,2010a; Hast等,2013; Cazanave等,2014)。
从可用文库中索引的大量化合物中,化合物LH601,苯磺酰基嘧啶酮2,N-苯基-苯磺酰胺和一系列1,4-二苯基-1,2,3-三唑可能是KEAP1抑制PPI非常适合的候选者(Hu等,2013; Jnoff等,2014; Bertrand等,2015; Wen等,2015; Nasiri等,2016)。这些研究详细描述了与KEAP1的原子相互作用、亲和力和结合的热力学参数。这些化合物的治疗功效将在未来的工作中进行分析,其中应该解决安全性,效力和血脑屏障渗透性。
表3. 选择的NRF2诱导剂充当NRF2-KEAP1蛋白质 - 蛋白质相互作用抑制剂
C. 用于NRF2激活的Keap1以外的药物靶
蛋白激酶GSK-3磷酸化NRF2的DSGIS序列中的两个丝氨酸残基以产生一个磷酸化依赖性降解基序或磷酸化二核(图2)。该磷酸二核被E3连接酶适配子β-TrCP识别,导致NRF2的泛素依赖性蛋白酶体降解。因此,GSK-3抑制剂应通过阻止这种磷酸二核的产生来阻止NRF2降解。GSK-3是AD和其他病理表型中的重要激酶。它使细胞骨架蛋白tau磷酸化,促进神经原纤维缠结的形成,神经原纤维缠结是病理性细胞内聚集体,其干扰轴突运输并导致神经元死亡(Silva等,2014)。因此,推测GSK-3抑制可能具有防止神经原纤维缠结形成和NRF2降解的双重益处。不幸的是,大多数用于开发GSK-3抑制剂的管道由于无效而已经停止,尽管在大多数情况下没有很好的证据表明靶标调节(Palomo和Martinez,2017)。从概念上讲,β-TrCP-磷酸化NRF2相互作用的抑制剂也应该导致NRF2活化,因为它们应该破坏NRF2降解的这一分支(图2)。β-TrCP的β-螺旋和含有NRF2磷酸二核的肽之间的分子相互作用已经通过NMR解决(Rada等,2012)。正如KEAP1/EGTE所发生的那样,最相关的氨基酸似乎是β-TrCP的几个精氨酸残基,其与DpSGIpS基序的两个磷酸丝氨酸相互作用。然而,能够抑制β-TrCP-磷酸化NRF2相互作用的小分子的发现仍在进行中。
已经开发了其他策略来抑制NRF2抑制因子BACH1,这是一种bZip蛋白,可与MAF蛋白形成异二聚体并阻断ARE基因的表达。已经在体外描述了HPP-4382化合物对BACH1的有效抑制(Attucks等,2014),但是,在完整的临床试验之前,必须在体内证明HPP-4382的安全性和功效特征。考虑到其他途径也可能影响NRF2的活性,我们有理由推测,组合方法将是激活该转录因子的最佳途径。
D. NRF2抑制剂
NRF2在构成性和高度过表达时具有与其致癌活性相关的“阴暗面”。因此,已提出NRF2抑制作为使癌细胞对化学治疗药物或放射疗法敏感的机制(Milkovic等,2017)。可以设想两种策略用小分子抑制NRF2:破坏NRF2和MAFs之间的bZip相互作用的PPI抑制剂,以及阻断NRF2-MAF与ARE结合的DNA-蛋白质相互作用抑制剂(图9C)。这两种策略都受到了阻碍,因为这类药物需要克服蛋白质-蛋白质之间以及蛋白质-DNA界面(在较小程度上)之间的大量自由能。 然而,已发现其他bZip转录因子如STAT3-STAT3,MYC-MAX和JUNFOS(Yap等,2011),并且正在描述NRF2-MAF的新的小分子。例如,malabaricone-A是一种促氧化合物,通过靶向NRF2克服了白血病抗性(Manna等,2015)。抗坏血酸(维生素C),一种著名的ROS清除剂,被发现通过降低NRF2/ARE复合物水平,降低GCLC基因的表达,降低GSH水平,来增强伊马替尼耐药癌细胞的敏感性(Tarumoto等,2004)。全反式维甲酸是NRF2抑制剂的另一个实例,其在体外和体内显着降低有效的亲电NRF2诱导物对NRF2的活化。它激活视黄酸受体α,其与NRF2形成复合物,因此阻碍转录因子与ARE基因的结合(Wang等,2007)。天然产物如鸦胆子苦醇(Ren等,2011; Olayanju等,2015),赭曲霉毒素A(Tarumoto等,2004; Limonciel和Jennings,2014)和葫芦巴碱(Arlt等,2013)也有被发现抑制NRF2。然而,他们的作用机制还没有完全明白。事实上,与现有化合物相关的一个重要问题是它们可能具有的深远的脱靶效应。例如,最近发现鸦胆子苦醇对蛋白质合成具有普遍和非特异性抑制作用,导致NRF2水平下降,而且许多其他快速转换蛋白下降,因此最近不鼓励使用鸦胆子苦醇(Harder et al。,2017))。类似地,兽医实践中使用的抗原虫剂卤虫酮通过抑制NRF2积累来增强癌细胞的化学敏感性,但这种效应似乎是间接的,它通过抑制核糖基转移RNA的合成,这是NRF2以及许多其他含脯氨酸的蛋白质的核糖体转化所强烈需要的(Tsuchida等,2017)。
最近报道了一种识别选择性NRF2抑制剂的新方法,即使用小分子抑制剂的高通量定量筛选(Singh等, 2016)。作者确定了一种名为ML385的一流化合物,它最有可能阻止NRF2与其他bZip共激活因子的结合。该化合物阻断NRF2转录活性并使KEAP1缺陷细胞对卡铂和其他化学治疗药物敏感。需要进一步的研究来确认ML385是否对NRF2具有选择性,或者它是否也抑制其他bZip转录因子。
鉴于NRF2在多种肿瘤病理表型中具有良好的全身效应,用小分子抑制剂特异性靶向NRF2似乎提供了一种良好的临床途径。然而,有必要确定用NRF2抑制剂治疗癌症是否会增加NRF2疾病组中的其他病理表型的风险。
E. 药物新用代替新药物的发现与开发
如前所述,许多化合物正在开发中,为与NRF2疾病组相关的病理表型提供益处。另一种方法是给已经在临床中用于某种病理机制的药物一个新的用途,用于治疗与NRF2有关的其他病理机制。本节根据一些常用药物在NRF2调控中的作用,为其重新定位提供依据。二甲双胍是T2DM的一线单药治疗。根据图6,它为与葡萄糖代谢相关的病理表型的NRF2亚群提供治疗益处。事实上,SFN可降低肝脏葡萄糖的产生,并改善T2DM患者的血糖控制(Axelsson等,2017)。有趣的是,一些证据表明,二甲双胍可能有效预防NRF2疾病组中的其他非糖尿病病理表型,包括心血管疾病(Nesti和Natali,2017),呼吸系统疾病(Sato等,2016),消化系统(Bauer和Duca,2016),神经退行性疾病(Markowicz-Piasecka等,2017),自身免疫(Schuiveling等,2017)和肿瘤(Heckman-Stoddard等,2017)疾病。二甲双胍的作用机制尚不完全清楚,但它涉及抑制线粒体复合物I,从而增加AMP/ATP比值(El-Mir等,2000; Owen等,2000)并导致能量传感器AMPK的激活(Hardie,2004; Rena等,2017)。重要的是,AMPK激活NRF2(Wang等,2017a; Zhao等,2017),并且该轴的药理学靶向可减轻中风后的炎症(Wang等,2017c)或内毒素暴露(Ci等,2017; Lv等,2017)。事实上,二甲双胍以AMPK依赖的方式激活NRF2,从而抑制临床前啮齿动物短暂性全脑缺血模型的炎症反应(Ashabi等, 2015;Kaisar等,2017)。葡萄糖代谢和炎症可能不是二甲双胍/NRF2作用的唯一病理机制。实际上,已经描述了氧化还原的其他有益效果(Kocer等,2014; Kelleni等,2015)和蛋白质稳态(Tsai等,2017)。
他汀类药物可预防和减少心血管病变表型。除了降脂作用外,他汀类药物似乎可以预防与NRF2网络相关的病理机制,如炎症(Pantan等,2016; Wu等,2016a; Hwang等,2017)和病理性ROS形成( Abdanipour等,2014)。它们是3-羟基-3-甲基-戊二酰-CoA还原酶的竞争性抑制剂,其催化胆固醇合成中的限速反应。其他多效性影响包括转录因子Krüppel样因子2的上调,其在肝硬化进展期间早期诱导,并减轻肝血管功能障碍的发展(Marrone等,2015)。最近的证据表明,至少一些他汀类药物会激活NRF2。在分离的肝细胞中进行的一项研究中,高浓度的辛伐他汀激活了NRF2,可能作为一种防御机制(Cho等,2013)。用洛伐他汀预处理神经干细胞激活NRF2途径并引发针对过氧化氢诱导的细胞死亡的保护(Abdanipour等,2014)。在肝硬化中,辛伐他汀激活由Krüppel样因子2和NRF2形成的轴,以减少星状细胞的氧化负荷和炎症反应,改善肝纤维化,内皮功能障碍和门静脉高压。辛伐他汀激活NRF2的机制尚不完全清楚,但似乎涉及NRF2相互作用组中发现的元素,如丝裂原活化蛋白激酶、PI3K/AKT途径(Jang等,2016)和GSK-3(Lin等,2016)。
从图4的NRF2相互作用组可以推断出药物新用的其他病例,特别是信号激酶。如图2C所示,GSK-3磷酸化NRF2的Neh6结构域,导致β-TrCP的识别和进一步泛素依赖性蛋白酶体降解。GSK-3在没有刺激的情况下是活跃的,而在激活AKT和其他激酶的信号级联导致GSK-3在其N端伪底物域磷酸化时是不活跃的。因此,已知靶向信号激酶的药物可用于上调(GSK-3抑制剂)或下调(PI3K/AKT抑制剂)NRF2特征。
GSK-3参与NRF2疾病组中发现的至少一些病理表型,例如糖尿病和神经退行性疾病(Beurel等,2015; Maqbool和Hoda,2017)。从天然和合成来源发现了广泛的GSK-3抑制剂(Khan等,2017),但将GSK-3抑制剂重新利用来增加NRF2活性的最佳证据可能源于临床使用锂作为情绪稳定剂(Chiu等,2013)。虽然此时在NRF2疾病组中未发现双相情感障碍和抑郁,但很明显它们表现出神经炎症和退行性病变表型,至少在小鼠模型中暗示NRF2失调(Martin-de-Saavedra等,2013; Freitas等,2016; Yao等,2016)。
NRF2相互作用组也为阻断信号激酶从而激活GSK-3的癌症药物抑制NRF2提供了一个理由。例如,表皮生长因子受体抑制剂厄洛替尼导致NRF2抑制,参与非小细胞肺癌中的肿瘤细胞感觉(Xiaobo等,2016)。用于治疗肝细胞癌的激酶级联抑制剂索拉非尼也导致NRF2及其下游靶标金属硫蛋白-1(Houessinon等,2016)和亚甲基四氢叶酸脱氢酶1的抑制(Lee等人,2017)。
最后,在两项相关研究中,迄今为止一直在寻找可能影响NRF2调节的再利用药物。使用基于荧光相关光谱的筛选系统,1633种药物中的两种显着增加HepG2细胞中的NRF2蛋白水平:叶绿酸和bonaphton(Yoshizaki等,2017)。在另一项研究中,分析了连接图数据库,其包含用1309种试剂处理的人细胞系的基因表达谱(Lamb等,2006; Iorio等,2010)(Zhang等,2017),通过激活NRF2以寻找有潜力的氧化还原调节剂(Xiong等,2014)。该研究发现阿司咪唑是一种有效的抗组胺药,用于过敏性疾病,是一种新型的NRF2激活剂。
VI. 生物标志物作为NRF2特征和参与监测目标
由于ROS的半衰期短,在毫秒或纳秒的范围内,对患者或群体研究中的氧化还原状态的评估受到阻碍(Ghezzi等,2017b)。因此,病理性ROS形成的生物标志物基于测量ROS留下的痕迹,ROS通常是细胞分子的末端氧化产物,其中许多是非特异性的(Frijhoff等,2015)。相反,NRF2的激活和其靶基因的后续表达是对生物体对病理性ROS形成的总暴露的间接但可靠的估计。由于NRF2激活是对环境应激源的一种成熟的细胞反应,所以它被认为是暴露于异生素的生物标志物。在肺部,通过对多个转录研究的数据挖掘,并结合独创性通路分析,报道了NRF2信号在健康吸烟者中上调,因此表明NRF2调控的抗氧化基因在预防烟草烟雾的毒性作用中发挥核心作用(Comandini等, 2010)。类似地,NRF2调控的一种酶NQO1在过量服用对乙酰氨基酚患者的肝组织中含量是正常水平的15倍(Aleksunes等, 2006)。疾病与营养之间的关联通常基于不可靠的自我报告(Archer等,2015)。通过激活NRF2来测量对营养素响应的生物标志物,据称具有有益效果,这可以提供一种可靠的方法来验证营养研究。但是,这种可能性仍未得到探索。与NRF2转录相关的变化可用作监测药物功效的生物标志物,所述药物旨在通过黄嘌呤氧化酶和NADPH氧化酶抑制剂减少病理性ROS形成。同样,可以通过定义全局蛋白和基因表达谱来检测和监测环境化学品的暴露情况(Ghezzi等,2017a)。这种方法类似于第一阶段药物代谢酶的使用,其中细胞色素P450是各种异种生物通过Ah受体诱导的,可以作为海洋污染的指标(Cajaraville等, 2000)。12周内每日口服富马酸酯与银屑病患者皮肤中NRF2靶基因表达增加有关(Onderdijk等,2014)。同样,在接受每日剂量CDDO-Me治疗3周的癌症患者外周血单核细胞中,NQO1的mRNA水平增加了5倍(Hong等, 2012)。
使用NRF2的转录特征作为生物标记需要很好地了解ARE基因活化所涉及的机制,因为大多数NRF2靶标受另外的转录因子调节。因此,分析几种ARE基因的表达很重要。例如,一项使用NRF2作为肺鳞癌治疗反应预测因子的研究提出使用28个基因来定义NRF2激活谱(Cescon等, 2015)。
VII. 结论
系统医学和网络药理学共同强调了NRF2在慢性疾病中起基础性作用的一组病理表型。这些疾病具有共同的机制,包括氧化,炎症和代谢改变。 本研究中提出的NRF2相互作用组,NRF2疾病组和NRF2药物组仍处于早期发展阶段,但它们代表了将NRF2构建为一种常见的治疗和系统医学方法的首次尝试。 即将完善的现有数据库和即将公布的临床结果数据将进一步提高这一新方法的准确性,这一新方法是以药理学和机制为基础的药物新用。本文为药物发现激活或抑制NRF2的综合策略提供了路线图,并强调需要为新药的开发或针对NRF2作为慢性病常见成分的药物的重新利用的转化而努力。Ⅷ. 参考文献
Abdanipour A, Tiraihi T, Noori-Zadeh A, Majdi A, and Gosaili R (2014) Evaluation of lovastatin effects on expression of anti-apoptotic Nrf2 and PGC-1a genes in neural stem cells treated with hydrogen peroxide. Mol Neurobiol 49:1364–1372.Abed DA, Goldstein M, Albanyan H, Jin H, and Hu L (2015) Discovery of direct inhibitors of Keap1-Nrf2 protein-protein interaction as potential therapeutic and preventive agents. Acta Pharm Sin B 5:285–299.
Aleksunes LM, Goedken M, and Manautou JE (2006) Up-regulation of NAD(P)H quinone oxidoreductase 1 during human liver injury. World J Gastroenterol 12: 1937–1940.
Aleksunes LM and Manautou JE (2007) Emerging role of Nrf2 in protecting against hepatic and gastrointestinal disease. Toxicol Pathol 35:459–473.
Al-Huseini LM, Aw Yeang HX, Sethu S, Alhumeed N, Hamdam JM, Tingle Y, Djouhri L, Kitteringham N, Park BK, Goldring CE, et al. (2013) Nuclear factorerythroid 2 (NF-E2) p45-related factor-2 (Nrf2) modulates dendritic cell immune function through regulation of p38 MAPK-cAMP-responsive element binding protein/activating transcription factor 1 signaling. J Biol Chem 288:22281–22288.
Al-Jaderi Z and Maghazachi AA (2015) Vitamin D3 and monomethyl fumarate enhance natural killer cell lysis of dendritic cells and ameliorate the clinical score in mice suffering from experimental autoimmune encephalomyelitis. Toxins (Basel) 7: 4730–4744.
Altmeyer PJ, Matthes U, Pawlak F, Hoffmann K, Frosch PJ, Ruppert P, Wassilew SW, Horn T, Kreysel HW, Lutz G, et al. (1994) Antipsoriatic effect of fumaric acid derivatives: results of a multicenter double-blind study in 100 patients. J Am Acad Dermatol 30:977–981.
Archer E, Pavela G, and Lavie CJ (2015) The inadmissibility of what we eat in America and NHANES dietary data in nutrition and obesity research and the scientific formulation of national dietary guidelines. Mayo Clin Proc 90:911–926.
Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, Fujita H, Hasegawa S, Takagi T, Wang FY, et al. (2007) The relationship between Helicobacter pylori infection and promoter polymorphism of the Nrf2 gene in chronic gastritis. Int J Mol Med 19:143–148.
Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, Fujita H, Yoshioka D, Okubo M, Hirata I, et al. (2008a) Nrf2 gene promoter polymorphism and gastric carcinogenesis. Hepatogastroenterology 55:750–754.
Arisawa T, Tahara T, Shibata T, Nagasaka M, Nakamura M, Kamiya Y, Fujita H, Yoshioka D, Okubo M, Sakata M, et al. (2008b) Nrf2 gene promoter polymorphism is associated with ulcerative colitis in a Japanese population. Hepatogastroenterology 55:394–397.
Arlt A, Sebens S, Krebs S, Geismann C, Grossmann M, Kruse ML, Schreiber S, and Schäfer H (2013) Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 32: 4825–4835.
Artaud-Macari E, Goven D, Brayer S, Hamimi A, Besnard V, Marchal-Somme J, Ali ZE, Crestani B, Kerdine-Römer S, Boutten A, et al. (2013) Nuclear factor erythroid 2-related factor 2 nuclear translocation induces myofibroblastic dedifferentiation in idiopathic pulmonary fibrosis. Antioxid Redox Signal 18:66–79.
Ashabi G, Khalaj L, Khodagholi F, Goudarzvand M, and Sarkaki A (2015) Pretreatment with metformin activates Nrf2 antioxidant pathways and inhibits inflammatory responses through induction of AMPK after transient global cerebral ischemia. Metab Brain Dis 30:747–754.
Attucks OC, Jasmer KJ, Hannink M, Kassis J, Zhong Z, Gupta S, Victory SF, Guzel M, Polisetti DR, Andrews R, et al. (2014) Induction of heme oxygenase I (HMOX1) by HPP-4382: a novel modulator of Bach1 activity. PLoS One 9:e101044.
Aviello G and Knaus UG (2017) ROS in gastrointestinal inflammation: rescue or sabotage? Br J Pharmacol 174:1704–1718.
Axelsson AS, Tubbs E, Mecham B, Chacko S, Nenonen HA, Tang Y, Fahey JW, Derry JMJ, Wollheim CB, Wierup N, et al. (2017) Sulforaphane reduces hepatic glucose production and improves glucose control in patients with type 2 diabetes. Sci Transl Med 9:1–12.
Bahadoran Z, Mirmiran P, Hosseinpanah F, Rajab A, Asghari G, and Azizi F (2012) Broccoli sprouts powder could improve serum triglyceride and oxidized LDL/LDLcholesterol ratio in type 2 diabetic patients: a randomized double-blind placebocontrolled clinical trial. Diabetes Res Clin Pract 96:348–354.
Baillie JK, Arner E, Daub C, De Hoon M, Itoh M, Kawaji H, Lassmann T, Carninci P, Forrest AR, Hayashizaki Y, et al.; FANTOM Consortium (2017) Analysis of the human monocyte-derived macrophage transcriptome and response to lipopolysaccharide provides new insights into genetic aetiology of inflammatory bowel disease. PLoS Genet 13:e1006641.
Baird L and Dinkova-Kostova AT (2013) Diffusion dynamics of the Keap1-Cullin3 interaction in single live cells. Biochem Biophys Res Commun 433:58–65. Banning A and Brigelius-Flohé R (2005) NF-kappaB, Nrf2, and HO-1 interplay in redox-regulated VCAM-1 expression. Antioxid Redox Signal 7:889–899.
Barabási AL, Gulbahce N, and Loscalzo J (2011) Network medicine: a network-based approach to human disease. Nat Rev Genet 12:56–68.
Batthyany CI and Lopez GV (2015) Nitroalkene Tocopherols and Analogs Thereof for Use in the Treatment and Prevention of Inflammation Related Conditions, Complexa, Radnor, PA. Bauer PV and Duca FA (2016) Targeting the gastrointestinal tract to treat type 2 diabetes. J Endocrinol 230:R95–R113.
Bergström P, von Otter M, Nilsson S, Nilsson AC, Nilsson M, Andersen PM, Hammarsten O, and Zetterberg H (2014) Association of NFE2L2 and KEAP1 haplotypes with amyotrophic lateral sclerosis. Amyotroph Lateral Scler Frontotemporal Degener 15:130–137.
Bertrand HC, Schaap M, Baird L, Georgakopoulos ND, Fowkes A, Thiollier C, Kachi H, Dinkova-Kostova AT, and Wells G (2015) Design, synthesis, and evaluation of triazole derivatives that induce Nrf2 dependent gene products and inhibit the Keap1-Nrf2 protein-protein interaction. J Med Chem 58:7186–7194.
Beurel E, Grieco SF, and Jope RS (2015) Glycogen synthase kinase-3 (GSK3): regulation, actions, and diseases. Pharmacol Ther 148:114–131. Bhatt JK, Thomas S, and Nanjan MJ (2012) Resveratrol supplementation improves glycemic control in type 2 diabetes mellitus. Nutr Res 32:537–541.
Blewett MM, Xie J, Zaro BW, Backus KM, Altman A, Teijaro JR, and Cravatt BF (2016) Chemical proteomic map of dimethyl fumarate-sensitive cysteines in primary human T cells. Sci Signal 9:rs10.
Bolaños JP (2016) Bioenergetics and redox adaptations of astrocytes to neuronal activity. J Neurochem 139 (Suppl 2):115–125.
Bourdonnay E, Morzadec C, Fardel O, and Vernhet L (2009) Redox-sensitive regulation of gene expression in human primary macrophages exposed to inorganic arsenic. J Cell Biochem 107:537–547.
Bowie A and O’Neill LA (2000) Oxidative stress and nuclear factor-kappaB activation: a reassessment of the evidence in the light of recent discoveries. Biochem Pharmacol 59:13–23.
Brasnyó P, Molnár GA, Mohás M, Markó L, Laczy B, Cseh J, Mikolás E, Szijártó IA, Mérei A, Halmai R, et al. (2011) Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br J Nutr 106:383–389.
Brennan MS, Matos MF, Richter KE, Li B, and Scannevin RH (2017) The NRF2 transcriptional target, OSGIN1, contributes to monomethyl fumarate-mediated cytoprotection in human astrocytes. Sci Rep 7:42054.
Brüne B, Dehne N, Grossmann N, Jung M, Namgaladze D, Schmid T, von Knethen A, and Weigert A (2013) Redox control of inflammation in macrophages. Antioxid Redox Signal 19:595–637.
Buendia I, Gómez-Rangel V, González-Lafuente L, Parada E, León R, Gameiro I, Michalska P, Laudon M, Egea J, and López MG (2015a) Neuroprotective mechanism of the novel melatonin derivative Neu-P11 in brain ischemia related models. Neuropharmacology 99:187–195.
Buendia I, Michalska P, Navarro E, Gameiro I, Egea J, and León R (2016) Nrf2-ARE pathway: an emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases. Pharmacol Ther 157:84–104.
Buendia I, Navarro E, Michalska P, Gameiro I, Egea J, Abril S, López A, GonzálezLafuente L, López MG, and León R (2015b) New melatonin-cinnamate hybrids as multi-target drugs for neurodegenerative diseases: Nrf2-induction, antioxidant effect and neuroprotection. Future Med Chem 7:1961–1969.
Cajaraville MP, Bebianno MJ, Blasco J, Porte C, Sarasquete C, and Viarengo A (2000) The use of biomarkers to assess the impact of pollution in coastal environments of the Iberian Peninsula: a practical approach. Sci Total Environ 247: 295–311.
Cazanave SC, Wang X, Zhou H, Rahmani M, Grant S, Durrant DE, Klaassen CD, Yamamoto M, and Sanyal AJ (2014) Degradation of Keap1 activates BH3-only proteins Bim and PUMA during hepatocyte lipoapoptosis. Cell Death Differ 21: 1303–1312.
Cescon DW, She D, Sakashita S, Zhu CQ, Pintilie M, Shepherd FA, and Tsao MS (2015) NRF2 pathway activation and adjuvant chemotherapy benefit in lung squamous cell carcinoma. Clin Cancer Res 21:2499–2505.
Chen H, Assmann JC, Krenz A, Rahman M, Grimm M, Karsten CM, Köhl J, Offermanns S, Wettschureck N, and Schwaninger M (2014) Hydroxycarboxylic acid receptor 2 mediates dimethyl fumarate’s protective effect in EAE. J Clin Invest 124:2188–2192.
Chen J, Yu Y, Ji T, Ma R, Chen M, Li G, Li F, Ding Q, Kang Q, Huang D, et al. (2016) Clinical implication of Keap1 and phosphorylated Nrf2 expression in hepatocellular carcinoma. Cancer Med 5:2678–2687.
Chen W, Sun Z, Wang XJ, Jiang T, Huang Z, Fang D, and Zhang DD (2009) Direct interaction between Nrf2 and p21(Cip1/WAF1) upregulates the Nrf2-mediated antioxidant response. Mol Cell 34:663–673.
Chen Y, Inoyama D, Kong AN, Beamer LJ, and Hu L (2011) Kinetic analyses of Keap1-Nrf2 interaction and determination of the minimal Nrf2 peptide sequence required for Keap1 binding using surface plasmon resonance.
Chem Biol Drug Des 78:1014–1021. Chen YC, Wu YR, Wu YC, Lee-Chen GJ, and Chen CM (2013) Genetic analysis of NFE2L2 promoter variation in Taiwanese Parkinson’s disease. Parkinsonism Relat Disord 19:247–250.
Chin MP, Reisman SA, Bakris GL, O’Grady M, Linde PG, McCullough PA, Packham D, Vaziri ND, Ward KW, Warnock DG, et al. (2014) Mechanisms contributing to adverse cardiovascular events in patients with type 2 diabetes mellitus and stage 4 chronic kidney disease treated with bardoxolone methyl. Am J Nephrol 39: 499–508.
Chiu CT, Wang Z, Hunsberger JG, and Chuang DM (2013) Therapeutic potential of mood stabilizers lithium and valproic acid: beyond bipolar disorder. Pharmacol Rev 65:105–142.
Cho HY and Kleeberger SR (2010) Nrf2 protects against airway disorders. Toxicol Appl Pharmacol 244:43–56.
Cho HY, Marzec J, and Kleeberger SR (2015) Functional polymorphisms in Nrf2: implications for human disease. Free Radic Biol Med 88 (Pt B):362–372.
Cho HY, Reddy SP, Yamamoto M, and Kleeberger SR (2004) The transcription factor NRF2 protects against pulmonary fibrosis. FASEB J 18:1258–1260.
Cho YE, Moon PG, Lee JE, Singh TS, Kang W, Lee HC, Lee MH, Kim SH, and Baek MC (2013) Integrative analysis of proteomic and transcriptomic data for identification of pathways related to simvastatin-induced hepatotoxicity. Proteomics 13: 1257–1275.
Chowdhry S, Zhang Y, McMahon M, Sutherland C, Cuadrado A, and Hayes JD (2013) Nrf2 is controlled by two distinct b-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 32:3765–3781.
Christou H, Morita T, Hsieh CM, Koike H, Arkonac B, Perrella MA, and Kourembanas S (2000) Prevention of hypoxia-induced pulmonary hypertension by enhancement of endogenous heme oxygenase-1 in the rat. Circ Res 86: 1224–1229.
Chuengsamarn S, Rattanamongkolgul S, Phonrat B, Tungtrongchitr R, and Jirawatnotai S (2014) Reduction of atherogenic risk in patients with type 2 diabetes by curcuminoid extract: a randomized controlled trial. J Nutr Biochem 25:144–150.
Ci X, Zhou J, Lv H, Yu Q, Peng L, and Hua S (2017) Betulin exhibits antiinflammatory activity in LPS-stimulated macrophages and endotoxin-shocked mice through an AMPK/AKT/Nrf2-dependent mechanism. Cell Death Dis 8:e2798.
Cleasby A, Yon J, Day PJ, Richardson C, Tickle IJ, Williams PA, Callahan JF, Carr R, Concha N, Kerns JK, et al. (2014) Structure of the BTB domain of Keap1 and its interaction with the triterpenoid antagonist CDDO. PLoS One 9:e98896.
Comandini A, Marzano V, Curradi G, Federici G, Urbani A, and Saltini C (2010) Markers of anti-oxidant response in tobacco smoke exposed subjects: a data-mining review. Pulm Pharmacol Ther 23:482–492.
Córdova EJ, Velázquez-Cruz R, Centeno F, Baca V, and Orozco L (2010) The NRF2 gene variant, -653G/A, is associated with nephritis in childhood-onset systemic lupus erythematosus. Lupus 19:1237–1242.
Croze E, Yamaguchi KD, Knappertz V, Reder AT, and Salamon H (2013) Interferonbeta-1b-induced short- and long-term signatures of treatment activity in multiple sclerosis. Pharmacogenomics J 13:443–451.
Cuadrado A (2015) Structural and functional characterization of Nrf2 degradation by glycogen synthase kinase 3/b-TrCP. Free Radic Biol Med 88 (Pt B):147–157.
Cuadrado A, Martín-Moldes Z, Ye J, and Lastres-Becker I (2014) Transcription factors NRF2 and NF-kB are coordinated effectors of the Rho family, GTP-binding protein RAC1 during inflammation. J Biol Chem 289:15244–15258.
Cuadrado A, Moreno-Murciano P, and Pedraza-Chaverri J (2009) The transcription factor Nrf2 as a new therapeutic target in Parkinson’s disease. Expert Opin Ther Targets 13:319–329.
Cui Y, Wang Q, Li X, and Zhang X (2013) Experimental nonalcoholic fatty liver disease in mice leads to cytochrome p450 2a5 upregulation through nuclear factor erythroid 2-like 2 translocation. Redox Biol 1:433–440.
Datta S, Kundu S, Ghosh P, De S, Ghosh A, and Chatterjee M (2014) Correlation of oxidant status with oxidative tissue damage in patients with rheumatoid arthritis. Clin Rheumatol 33:1557–1564.
de la Vega MR, Dodson M, Gross C, Manzour H, Lantz RC, Chapman E, Wang T, Black SM, Garcia JG, and Zhang DD (2016) Role of Nrf2 and autophagy in acute lung injury. Curr Pharmacol Rep 2:91–101.
DeNicola GM, Karreth FA, Humpton TJ, Gopinathan A, Wei C, Frese K, Mangal D, Yu KH, Yeo CJ, Calhoun ES, et al. (2011) Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature 475:106–109.
de Zeeuw D, Akizawa T, Audhya P, Bakris GL, Chin M, Christ-Schmidt H, Goldsberry A, Houser M, Krauth M, Lambers Heerspink HJ, et al.; BEACON Trial Investigators (2013) Bardoxolone methyl in type 2 diabetes and stage 4 chronic kidney disease. N Engl J Med 369:2492–2503.
Diotallevi M, Checconi P, Palamara AT, Celestino I, Coppo L, Holmgren A, Abbas K, Peyrot F, Mengozzi M, and Ghezzi P (2017) Glutathione fine-tunes the innate immune response toward antiviral pathways in a macrophage cell line independently of its antioxidant properties. Front Immunol 8:1239.
Duran CG, Burbank AJ, Mills KH, Duckworth HR, Aleman MM, Kesic MJ, Peden DB, Pan Y, Zhou H, and Hernandez ML (2016) A proof-of-concept clinical study examining the NRF2 activator sulforaphane against neutrophilic airway inflammation. Respir Res 17:89.
Eades G, Yang M, Yao Y, Zhang Y, and Zhou Q (2011) miR-200a regulates Nrf2 activation by targeting Keap1 mRNA in breast cancer cells. J Biol Chem 286: 40725–40733.
Egea J, Buendia I, Parada E, Navarro E, Rada P, Cuadrado A, López MG, García AG, and León R (2015) Melatonin-sulforaphane hybrid ITH12674 induces neuroprotection in oxidative stress conditions by a ‘drug-prodrug’ mechanism of action. Br J Pharmacol 172:1807–1821.
Ellrichmann G, Petrasch-Parwez E, Lee DH, Reick C, Arning L, Saft C, Gold R, and Linker RA (2011) Efficacy of fumaric acid esters in the R6/2 and YAC128 models of Huntington’s disease. PLoS One 6:e16172.
El-Mir MY, Nogueira V, Fontaine E, Avéret N, Rigoulet M, and Leverve X (2000) Dimethylbiguanide inhibits cell respiration via an indirect effect targeted on the respiratory chain complex I. J Biol Chem 275:223–228.
Eminel S, Jin N, Rostami M, Dibbert S, Mrowietz U, and Suhrkamp I (2017) Dimethyl- and monomethylfumarate regulate indoleamine 2,3-dioxygenase (IDO) activity in human immune cells. Exp Dermatol 26:685–690.
Emmink BL, Verheem A, Van Houdt WJ, Steller EJ, Govaert KM, Pham TV, Piersma SR, Borel Rinkes IH, Jimenez CR, and Kranenburg O (2013) The secretome of colon cancer stem cells contains drug-metabolizing enzymes. J Proteomics 91:84–96.
Evans MD, Cooke MS, Akil M, Samanta A, and Lunec J (2000) Aberrant processing of oxidative DNA damage in systemic lupus erythematosus. Biochem Biophys Res Commun 273:894–898.
Fagone P, Patti F, Mangano K, Mammana S, Coco M, Touil-Boukoffa C, Chikovani T, Di Marco R, and Nicoletti F (2013) Heme oxygenase-1 expression in peripheral blood mononuclear cells correlates with disease activity in multiple sclerosis. J Neuroimmunol 261:82–86.
Finkelstein R, Fraser RS, Ghezzo H, and Cosio MG (1995) Alveolar inflammation and its relation to emphysema in smokers. Am J Respir Crit Care Med 152:1666–1672.
Fox RJ, Miller DH, Phillips JT, Hutchinson M, Havrdova E, Kita M, Yang M, Raghupathi K, Novas M, Sweetser MT, et al.; CONFIRM Study Investigators (2012) Placebo-controlled phase 3 study of oral BG-12 or glatiramer in multiple sclerosis. N Engl J Med 367:1087–1097.
Freitas AE, Egea J, Buendia I, Gómez-Rangel V, Parada E, Navarro E, Casas AI, Wojnicz A, Ortiz JA, Cuadrado A, et al. (2016) Agmatine, by improving neuroplasticity markers and inducing Nrf2, prevents corticosterone-induced depressivelike behavior in mice. Mol Neurobiol 53:3030–3045.
Frijhoff J, Winyard PG, Zarkovic N, Davies SS, Stocker R, Cheng D, Knight AR, Taylor EL, Oettrich J, Ruskovska T, et al. (2015) Clinical relevance of biomarkers of oxidative stress. Antioxid Redox Signal 23:1144–1170.
Frostegård J, Svenungsson E, Wu R, Gunnarsson I, Lundberg IE, Klareskog L, Hörkkö S, and Witztum JL (2005) Lipid peroxidation is enhanced in patients with systemic lupus erythematosus and is associated with arterial and renal disease manifestations. Arthritis Rheum 52:192–200.
Gao J, Chang MT, Johnsen HC, Gao SP, Sylvester BE, Sumer SO, Zhang H, Solit DB, Taylor BS, Schultz N, et al. (2017) 3D clusters of somatic mutations in cancer reveal numerous rare mutations as functional targets. Genome Med 9:4.
Ghanim H, Sia CL, Korzeniewski K, Lohano T, Abuaysheh S, Marumganti A, Chaudhuri A, and Dandona P (2011) A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, highcarbohydrate meal. J Clin Endocrinol Metab 96:1409–1414.
Ghezzi P, Floridi L, Boraschi D, Cuadrado A, Manda G, Levic S, D’Acquisto F, Hamilton A, Athersuch TJ, and Selley L (2017a) Oxidative stress and inflammation induced by environmental and psychological stressors: a biomarker perspective. Antioxid Redox Signal [published ahead of print].
Ghezzi P, Jaquet V, Marcucci F, and Schmidt HHHW (2017b) The oxidative stress theory of disease: levels of evidence and epistemological aspects. Br J Pharmacol 174:1784–1796.
Ghoreschi K, Brück J, Kellerer C, Deng C, Peng H, Rothfuss O, Hussain RZ, Gocke AR, Respa A, Glocova I, et al. (2011) Fumarates improve psoriasis and multiple sclerosis by inducing type II dendritic cells. J Exp Med 208:2291–2303.
Gillard GO, Collette B, Anderson J, Chao J, Scannevin RH, Huss DJ, and Fontenot JD (2015) DMF, but not other fumarates, inhibits NF-kB activity in vitro in an Nrf2-independent manner. J Neuroimmunol 283:74–85.
Goh KI, Cusick ME, Valle D, Childs B, Vidal M, and Barabási AL (2007) The human disease network. Proc Natl Acad Sci USA 104:8685–8690.
Gold R, Kappos L, Arnold DL, Bar-Or A, Giovannoni G, Selmaj K, Tornatore C, Sweetser MT, Yang M, Sheikh SI, et al.; DEFINE Study Investigators (2012) Placebo-controlled phase 3 study of oral BG-12 for relapsing multiple sclerosis. N Engl J Med 367:1098–1107.
Gonzalez-Donquiles C, Alonso-Molero J, Fernandez-Villa T, Vilorio-Marqués L, Molina AJ, and Martín V (2017) The NRF2 transcription factor plays a dual role in colorectal cancer: a systematic review. PLoS One 12:e0177549.
Goven D, Boutten A, Leçon-Malas V, Marchal-Sommé J, Amara N, Crestani B, Fournier M, Lesèche G, Soler P, Boczkowski J, et al. (2008) Altered Nrf2/Keap1- Bach1 equilibrium in pulmonary emphysema. Thorax 63:916–924.
Griendling KK and FitzGerald GA (2003a) Oxidative stress and cardiovascular injury: part I: basic mechanisms and in vivo monitoring of ROS. Circulation 108: 1912–1916.
Griendling KK and FitzGerald GA (2003b) Oxidative stress and cardiovascular injury: part II: animal and human studies. Circulation 108:2034–2040.
Guan CP, Zhou MN, Xu AE, Kang KF, Liu JF, Wei XD, Li YW, Zhao DK, and Hong WS (2008) The susceptibility to vitiligo is associated with NF-E2-related factor2 (Nrf2) gene polymorphisms: a study on Chinese Han population. Exp Dermatol 17: 1059–1062.
Guney E, Menche J, Vidal M, and Barábasi AL (2016) Network-based in silico drug efficacy screening. Nat Commun 7:10331.
Guney E and Oliva B (2014) Analysis of the robustness of network-based diseasegene prioritization methods reveals redundancy in the human interactome and functional diversity of disease-genes. PLoS One 9:e94686.
Han SJ, Ahn Y, Park I, Zhang Y, Kim I, Kim HW, Ku CS, Chay KO, Yang SY, Ahn BW, et al. (2015) Assay of the redox state of the tumor suppressor PTEN by mobility shift. Methods 77–78:58–62.
Hancock R, Schaap M, Pfister H, and Wells G (2013) Peptide inhibitors of the Keap1- Nrf2 protein-protein interaction with improved binding and cellular activity. Org Biomol Chem 11:3553–3557.
Harder B, Tian W, La Clair JJ, Tan AC, Ooi A, Chapman E, and Zhang DD (2017) Brusatol overcomes chemoresistance through inhibition of protein translation. Mol Carcinog 56:1493–1500.
Hardie DG (2004) The AMP-activated protein kinase pathway–new players upstream and downstream. J Cell Sci 117:5479–5487.
Harrison D, Griendling KK, Landmesser U, Hornig B, and Drexler H (2003) Role of oxidative stress in atherosclerosis. Am J Cardiol 91:7A–11A.
Hart PC, Ratti BA, Mao M, Ansenberger-Fricano K, Shajahan-Haq AN, Tyner AL, Minshall RD, and Bonini MG (2016) Caveolin-1 regulates cancer cell metabolism via scavenging Nrf2 and suppressing MnSOD-driven glycolysis. Oncotarget 7: 308–322.
Harvey CJ, Thimmulappa RK, Sethi S, Kong X, Yarmus L, Brown RH, Feller-Kopman D, Wise R, and Biswal S (2011) Targeting Nrf2 signaling improves bacterial clearance by alveolar macrophages in patients with COPD and in a mouse model. Sci Transl Med 3:78ra32.
Hast BE, Goldfarb D, Mulvaney KM, Hast MA, Siesser PF, Yan F, Hayes DN, and Major MB (2013) Proteomic analysis of ubiquitin ligase KEAP1 reveals associated proteins that inhibit NRF2 ubiquitination. Cancer Res 73:2199–2210.
Hayes JD and Dinkova-Kostova AT (2014) The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem Sci 39: 199–218.
Heckman-Stoddard BM, DeCensi A, Sahasrabuddhe VV, and Ford LG (2017) Repurposing metformin for the prevention of cancer and cancer recurrence. Diabetologia 60:1639–1647.
Hidalgo CA, Blumm N, Barabási AL, and Christakis NA (2009) A dynamic network approach for the study of human phenotypes. PLOS Comput Biol 5:e1000353.
Hong DS, Kurzrock R, Supko JG, He X, Naing A, Wheler J, Lawrence D, Eder JP, Meyer CJ, Ferguson DA, et al. (2012) A phase I first-in-human trial of bardoxolone methyl in patients with advanced solid tumors and lymphomas. Clin Cancer Res 18:3396–3406.
Hopkins PN, Wu LL, Hunt SC, James BC, Vincent GM, and Williams RR (1996) Higher serum bilirubin is associated with decreased risk for early familial coronary artery disease. Arterioscler Thromb Vasc Biol 16:250–255.
Houessinon A, François C, Sauzay C, Louandre C, Mongelard G, Godin C, Bodeau S, Takahashi S, Saidak Z, Gutierrez L, et al. (2016) Metallothionein-1 as a biomarker of altered redox metabolism in hepatocellular carcinoma cells exposed to sorafenib. Mol Cancer 15:38.
Houghton CA, Fassett RG, and Coombes JS (2016) Sulforaphane and other nutrigenomic Nrf2 activators: can the clinician’s expectation be matched by the reality? Oxid Med Cell Longev 2016:7857186.
Hu L, Magesh S, Chen L, Wang L, Lewis TA, Chen Y, Khodier C, Inoyama D, Beamer LJ, Emge TJ, et al. (2013) Discovery of a small-molecule inhibitor and cellular probe of Keap1-Nrf2 protein-protein interaction. Bioorg Med Chem Lett 23: 3039–3043.
Hua CC, Chang LC, Tseng JC, Chu CM, Liu YC, and Shieh WB (2010) Functional haplotypes in the promoter region of transcription factor Nrf2 in chronic obstructive pulmonary disease. Dis Markers 28:185–193.
Hur W and Gray NS (2011) Small molecule modulators of antioxidant response pathway. Curr Opin Chem Biol 15:162–173.
Hwang AR, Han JH, Lim JH, Kang YJ, and Woo CH (2017) Fluvastatin inhibits AGE-induced cell proliferation and migration via an ERK5-dependent Nrf2 pathway in vascular smooth muscle cells. PLoS One 12:e0178278.
Innamorato NG, Rojo AI, García-Yagüe AJ, Yamamoto M, de Ceballos ML, and Cuadrado A (2008) The transcription factor Nrf2 is a therapeutic target against brain inflammation. J Immunol 181:680–689.
Inoyama D, Chen Y, Huang X, Beamer LJ, Kong AN, and Hu L (2012) Optimization of fluorescently labeled Nrf2 peptide probes and the development of a fluorescence polarization assay for the discovery of inhibitors of Keap1-Nrf2 interaction. J Biomol Screen 17:435–447.
Iorio F, Isacchi A, di Bernardo D, and Brunetti-Pierri N (2010) Identification of small molecules enhancing autophagic function from drug network analysis. Autophagy 6:1204–1205.
Ishii T and Mann GE (2014) Redox status in mammalian cells and stem cells during culture in vitro: critical roles of Nrf2 and cystine transporter activity in the maintenance of redox balance. Redox Biol 2:786–794.
Jang HJ, Hong EM, Kim M, Kim JH, Jang J, Park SW, Byun HW, Koh DH, Choi MH, Kae SH, et al. (2016) Simvastatin induces heme oxygenase-1 via NF-E2-related factor 2 (Nrf2) activation through ERK and PI3K/Akt pathway in colon cancer. Oncotarget 7:46219–46229.
Jansen T, Hortmann M, Oelze M, Opitz B, Steven S, Schell R, Knorr M, Karbach S, Schuhmacher S, Wenzel P, et al. (2010) Conversion of biliverdin to bilirubin by biliverdin reductase contributes to endothelial cell protection by heme oxygenase1-evidence for direct and indirect antioxidant actions of bilirubin. J Mol Cell Cardiol 49:186–195.
Jay D, Hitomi H, and Griendling KK (2006) Oxidative stress and diabetic cardiovascular complications. Free Radic Biol Med 40:183–192.
Jazwa A, Rojo AI, Innamorato NG, Hesse M, Fernández-Ruiz J, and Cuadrado A (2011) Pharmacological targeting of the transcription factor Nrf2 at the basal ganglia provides disease modifying therapy for experimental parkinsonism. Antioxid Redox Signal 14:2347–2360.
Jiang T, Huang Z, Lin Y, Zhang Z, Fang D, and Zhang DD (2010) The protective role of Nrf2 in streptozotocin-induced diabetic nephropathy. Diabetes 59:850–860.
Jiang T, Tian F, Zheng H, Whitman SA, Lin Y, Zhang Z, Zhang N, and Zhang DD (2014a) Nrf2 suppresses lupus nephritis through inhibition of oxidative injury and the NF-kB-mediated inflammatory response. Kidney Int 85:333–343.
Jiang ZY, Lu MC, Xu LL, Yang TT, Xi MY, Xu XL, Guo XK, Zhang XJ, You QD, and Sun HP (2014b) Discovery of potent Keap1-Nrf2 protein-protein interaction inhibitor based on molecular binding determinants analysis [published correction appears in J Med Chem (2014) 57:4406]. J Med Chem 57: 2736–2745.
Jiang ZY, Lu MC, and You QD (2016) Discovery and development of Kelch-like ECHassociated protein 1: Nuclear factor erythroid 2-related factor 2 (KEAP1:NRF2) protein-protein interaction inhibitors: achievements, challenges, and future directions. J Med Chem 59:10837–10858.
Jiménez-Osorio AS, González-Reyes S, García-Niño WR, Moreno-Macías H, Rodríguez-Arellano ME, Vargas-Alarcón G, Zúñiga J, Barquera R, Pedraza-Chaverri J, Meza-Espinoza JP, et al. (2017) Corrigendum to “association of nuclear factorerythroid 2-related factor 2, thioredoxin interacting protein, and heme oxygenase-1 gene polymorphisms with diabetes and obesity in Mexican patients.” Oxid Med Cell Longev 2017:7543194.
Jiménez-Osorio AS, Picazo A, González-Reyes S, Barrera-Oviedo D, Rodríguez-Arellano ME, and Pedraza-Chaverri J (2014) Nrf2 and redox status in prediabetic and diabetic patients. Int J Mol Sci 15:20290–20305.
Jnoff E, Albrecht C, Barker JJ, Barker O, Beaumont E, Bromidge S, Brookfield F, Brooks M, Bubert C, Ceska T, et al. (2014) Binding mode and structure-activity relationships around direct inhibitors of the Nrf2-Keap1 complex. ChemMedChem 9:699–705.
Jo C, Gundemir S, Pritchard S, Jin YN, Rahman I, and Johnson GV (2014) Nrf2 reduces levels of phosphorylated tau protein by inducing autophagy adaptor protein NDP52. Nat Commun 5:3496.
Johnson DA, Amirahmadi S, Ward C, Fabry Z, and Johnson JA (2010) The absence of the pro-antioxidant transcription factor Nrf2 exacerbates experimental autoimmune encephalomyelitis. Toxicol Sci 114:237–246.
Johnson DA and Johnson JA (2015) Nrf2–a therapeutic target for the treatment of neurodegenerative diseases. Free Radic Biol Med 88 (Pt B):253–267.
Johnson NM, Egner PA, Baxter VK, Sporn MB, Wible RS, Sutter TR, Groopman JD, Kensler TW, and Roebuck BD (2014) Complete protection against aflatoxin B(1)- induced liver cancer with a triterpenoid: DNA adduct dosimetry, molecular signature, and genotoxicity threshold. Cancer Prev Res (Phila) 7:658–665.
Jones RM, Desai C, Darby TM, Luo L, Wolfarth AA, Scharer CD, Ardita CS, Reedy AR, Keebaugh ES, and Neish AS (2015) Lactobacilli modulate epithelial cytoprotection through the Nrf2 pathway. Cell Reports 12:1217–1225.
Jung KA, Lee S, and Kwak MK (2017) NFE2L2/NRF2 activity is linked to mitochondria and AMP-activated protein kinase signaling in cancers through miR181c/mitochondria-encoded cytochrome c oxidase regulation. Antioxid Redox Signal 27:945–961.
Kaidery NA, Banerjee R, Yang L, Smirnova NA, Hushpulian DM, Liby KT, Williams CR, Yamamoto M, Kensler TW, Ratan RR, et al. (2013) Targeting Nrf2-mediated gene transcription by extremely potent synthetic triterpenoids attenuates dopaminergic neurotoxicity in the MPTP mouse model of Parkinson’s disease. Antioxid Redox Signal 18:139–157.
Kaisar MA, Villalba H, Prasad S, Liles T, Sifat AE, Sajja RK, Abbruscato TJ, and Cucullo L (2017) Offsetting the impact of smoking and e-cigarette vaping on the cerebrovascular system and stroke injury: is Metformin a viable countermeasure? Redox Biol 13:353–362.
Kastrati I, Siklos MI, Calderon-Gierszal EL, El-Shennawy L, Georgieva G, Thayer EN, Thatcher GR, and Frasor J (2016) Dimethyl fumarate inhibits the nuclear factor kB pathway in breast cancer cells by covalent modification of p65 protein. J Biol Chem 291:3639–3647.
Kelleni MT, Amin EF, and Abdelrahman AM (2015) Effect of metformin and sitagliptin on doxorubicin-induced cardiotoxicity in rats: impact of oxidative stress, inflammation, and apoptosis. J Toxicol 2015:424813. Kensler TW, Egner PA, Agyeman AS, Visvanathan K, Groopman JD, Chen JG, Chen TY, Fahey JW, and Talalay P (2013) Keap1-nrf2 signaling: a target for cancer prevention by sulforaphane. Top Curr Chem 329:163–177.
Khan I, Tantray MA, Alam MS, and Hamid H (2017) Natural and synthetic bioactive inhibitors of glycogen synthase kinase. Eur J Med Chem 125:464–477. Kim JE, You DJ, Lee C, Ahn C, Seong JY, and Hwang JI (2010a) Suppression of NF-kappaB signaling by KEAP1 regulation of IKKbeta activity through autophagic degradation and inhibition of phosphorylation. Cell Signal 22:1645–1654.
Kim T, Kim YJ, Han IH, Lee D, Ham J, Kang KS, and Lee JW (2015) The synthesis of sulforaphane analogues and their protection effect against cisplatin induced cytotoxicity in kidney cells. Bioorg Med Chem Lett 25:62–66.
Kim YR, Oh JE, Kim MS, Kang MR, Park SW, Han JY, Eom HS, Yoo NJ, and Lee SH (2010b) Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin. J Pathol 220:446–451.
Kitagishi Y and Matsuda S (2013) Redox regulation of tumor suppressor PTEN in cancer and aging (Review). Int J Mol Med 31:511–515.
Kitsak M, Sharma A, Menche J, Guney E, Ghiassian SD, Loscalzo J, and Barabási AL (2016) Tissue specificity of human disease module. Sci Rep 6:35241. Kobayashi EH, Suzuki T, Funayama R, Nagashima T, Hayashi M, Sekine H, Tanaka N, Moriguchi T, Motohashi H, Nakayama K, et al. (2016) Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nat Commun 7:11624.
Kocer D, Bayram F, and Diri H (2014) The effects of metformin on endothelial dysfunction, lipid metabolism and oxidative stress in women with polycystic ovary syndrome. Gynecol Endocrinol 30:367–371.
Komatsu M, Kurokawa H, Waguri S, Taguchi K, Kobayashi A, Ichimura Y, Sou YS, Ueno I, Sakamoto A, Tong KI, et al. (2010) The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat Cell Biol 12:213–223.
Kong X, Thimmulappa R, Craciun F, Harvey C, Singh A, Kombairaju P, Reddy SP, Remick D, and Biswal S (2011) Enhancing Nrf2 pathway by disruption of Keap1 in myeloid leukocytes protects against sepsis. Am J Respir Crit Care Med 184: 928–938.
Kruger AL, Peterson SJ, Schwartzman ML, Fusco H, McClung JA, Weiss M, Shenouda S, Goodman AI, Goligorsky MS, Kappas A, et al. (2006) Up-regulation of heme oxygenase provides vascular protection in an animal model of diabetes through its antioxidant and antiapoptotic effects. J Pharmacol Exp Ther 319: 1144–1152.
Kruse ML, Friedrich M, Arlt A, Röcken C, Egberts JH, Sebens S, and Schäfer H (2016) Colonic lamina propria inflammatory cells from patients with IBD induce the nuclear factor-E2 related factor-2 thereby leading to greater proteasome activity and apoptosis protection in human colonocytes. Inflamm Bowel Dis 22: 2593–2606.
Kuang X, Scofield VL, Yan M, Stoica G, Liu N, and Wong PK (2009) Attenuation of oxidative stress, inflammation and apoptosis by minocycline prevents retrovirusinduced neurodegeneration in mice. Brain Res 1286:174–184.
Kurinna S and Werner S (2015) NRF2 and microRNAs: new but awaited relations. Biochem Soc Trans 43:595–601.
Kwak JY, Takeshige K, Cheung BS, and Minakami S (1991) Bilirubin inhibits the activation of superoxide-producing NADPH oxidase in a neutrophil cell-free system. Biochim Biophys Acta 1076:369–373.
Lamb J, Crawford ED, Peck D, Modell JW, Blat IC, Wrobel MJ, Lerner J, Brunet JP, Subramanian A, Ross KN, et al. (2006) The connectivity map: using geneexpression signatures to connect small molecules, genes, and disease. Science 313: 1929–1935.
Lastres-Becker I, García-Yagüe AJ, Scannevin RH, Casarejos MJ, Kügler S, Rábano A, and Cuadrado A (2016) Repurposing the NRF2 activator dimethyl fumarate as therapy against synucleinopathy in Parkinson’s disease. Antioxid Redox Signal 25: 61–77.
Lastres-Becker I, Innamorato NG, Jaworski T, Rábano A, Kügler S, Van Leuven F, and Cuadrado A (2014) Fractalkine activates NRF2/NFE2L2 and heme oxygenase 1 to restrain tauopathy-induced microgliosis. Brain 137:78–91.
Lastres-Becker I, Ulusoy A, Innamorato NG, Sahin G, Rábano A, Kirik D, and Cuadrado A (2012) a-Synuclein expression and Nrf2 deficiency cooperate to aggravate protein aggregation, neuronal death and inflammation in early-stage Parkinson’s disease. Hum Mol Genet 21:3173–3192.
Lee D, Xu IM, Chiu DK, Lai RK, Tse AP, Lan Li L, Law CT, Tsang FH, Wei LL, Chan CY, et al. (2017) Folate cycle enzyme MTHFD1L confers metabolic advantages in hepatocellular carcinoma. J Clin Invest 127:1856–1872.
Lee DF, Kuo HP, Liu M, Chou CK, Xia W, Du Y, Shen J, Chen CT, Huo L, Hsu MC, et al. (2009) KEAP1 E3 ligase-mediated downregulation of NF-kappaB signaling by targeting IKKbeta. Mol Cell 36:131–140.
Lee SR, Yang KS, Kwon J, Lee C, Jeong W, and Rhee SG (2002) Reversible inactivation of the tumor suppressor PTEN by H2O2. J Biol Chem 277:20336–20342.
Li B, Cui W, Liu J, Li R, Liu Q, Xie XH, Ge XL, Zhang J, Song XJ, Wang Y, et al. (2013) Sulforaphane ameliorates the development of experimental autoimmune encephalomyelitis by antagonizing oxidative stress and Th17-related inflammation in mice. Exp Neurol 250:239–249.
Li J, Stein TD, and Johnson JA (2004) Genetic dissection of systemic autoimmune disease in Nrf2-deficient mice. Physiol Genomics 18:261–272.
Licht-Mayer S, Wimmer I, Traffehn S, Metz I, Brück W, Bauer J, Bradl M, and Lassmann H (2015) Cell type-specific Nrf2 expression in multiple sclerosis lesions. Acta Neuropathol 130:263–277.
Limonciel A and Jennings P (2014) A review of the evidence that ochratoxin A is an Nrf2 inhibitor: implications for nephrotoxicity and renal carcinogenicity. Toxins (Basel) 6:371–379.
Lin CH, Lin HI, Chen ML, Lai TT, Cao LP, Farrer MJ, Wu RM, and Chien CT (2016) Lovastatin protects neurite degeneration in LRRK2-G2019S parkinsonism through activating the Akt/Nrf pathway and inhibiting GSK3b activity. Hum Mol Genet 25: 1965–1978.
Lin SX, Lisi L, Dello Russo C, Polak PE, Sharp A, Weinberg G, Kalinin S, and Feinstein DL (2011) The anti-inflammatory effects of dimethyl fumarate in astrocytes involve glutathione and haem oxygenase-1. ASN Neuro 3:75–84.
Lin W, Wu RT, Wu T, Khor TO, Wang H, and Kong AN (2008) Sulforaphane suppressed LPS-induced inflammation in mouse peritoneal macrophages through Nrf2 dependent pathway. Biochem Pharmacol 76:967–973.
Linker RA, Lee DH, Ryan S, van Dam AM, Conrad R, Bista P, Zeng W, Hronowsky X, Buko A, Chollate S, et al. (2011) Fumaric acid esters exert neuroprotective effects in neuroinflammation via activation of the Nrf2 antioxidant pathway. Brain 134: 678–692.
Liu GH, Qu J, and Shen X (2008) NF-kappaB/p65 antagonizes Nrf2-ARE pathway by depriving CBP from Nrf2 and facilitating recruitment of HDAC3 to MafK. Biochim Biophys Acta 1783:713–727.
Liu TS, Pei YH, Peng YP, Chen J, Jiang SS, and Gong JB (2014) Oscillating high glucose enhances oxidative stress and apoptosis in human coronary artery endothelial cells. J Endocrinol Invest 37:645–651.
Lo SC, Li X, Henzl MT, Beamer LJ, and Hannink M (2006) Structure of the Keap1: Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J 25: 3605–3617.
LoGerfo A, Chico L, Borgia L, Petrozzi L, Rocchi A, D’Amelio A, Carlesi C, Caldarazzo Ienco E, Mancuso M, and Siciliano G (2014) Lack of association between nuclear factor erythroid-derived 2-like 2 promoter gene polymorphisms and oxidative stress biomarkers in amyotrophic lateral sclerosis patients. Oxid Med Cell Longev 2014:432626.
Lu K, Alcivar AL, Ma J, Foo TK, Zywea S, Mahdi A, Huo Y, Kensler TW, Gatza ML, and Xia B (2017) NRF2 induction supporting breast cancer cell survival is enabled by oxidative stress-induced DPP3-KEAP1 interaction. Cancer Res 77: 2881–2892.
Lu MC, Jiao Q, Liu T, Tan SJ, Zhou HS, You QD, and Jiang ZY (2018) Discovery of a head-to-tail cyclic peptide as the Keap1-Nrf2 protein-protein interaction inhibitor with high cell potency. Eur J Med Chem 143:1578–1589.
Lv H, Liu Q, Wen Z, Feng H, Deng X, and Ci X (2017) Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute lung injury via induction of AMPK/GSK3bNrf2 signal axis. Redox Biol 12:311–324.
Ma Q (2013) Role of nrf2 in oxidative stress and toxicity. Annu Rev Pharmacol Toxicol 53:401–426.
Ma Q, Battelli L, and Hubbs AF (2006) Multiorgan autoimmune inflammation, enhanced lymphoproliferation, and impaired homeostasis of reactive oxygen species in mice lacking the antioxidant-activated transcription factor Nrf2. Am J Pathol 168:1960–1974.
Maicas N, Ferrándiz ML, Brines R, Ibáñez L, Cuadrado A, Koenders MI, van den Berg WB, and Alcaraz MJ (2011) Deficiency of Nrf2 accelerates the effector phase of arthritis and aggravates joint disease. Antioxid Redox Signal 15:889–901.
Manna A, De Sarkar S, De S, Bauri AK, Chattopadhyay S, and Chatterjee M (2015) The variable chemotherapeutic response of malabaricone-A in leukemic and solid tumor cell lines depends on the degree of redox imbalance. Phytomedicine 22: 713–723.
Maqbool M and Hoda N (2017) GSK3 inhibitors in the therapeutic development of diabetes, cancer and neurodegeneration: past, present and future. Curr Pharm Des 23:4332–4350.
Marcotte D, Zeng W, Hus JC, McKenzie A, Hession C, Jin P, Bergeron C, Lugovskoy A, Enyedy I, Cuervo H, et al. (2013) Small molecules inhibit the interaction of Nrf2 and the Keap1 Kelch domain through a non-covalent mechanism. Bioorg Med Chem 21:4011–4019.
Markart P, Luboeinski T, Korfei M, Schmidt R, Wygrecka M, Mahavadi P, Mayer K, Wilhelm J, Seeger W, Guenther A, et al. (2009) Alveolar oxidative stress is associated with elevated levels of nonenzymatic low-molecular-weight antioxidants in patients with different forms of chronic fibrosing interstitial lung diseases. Antioxid Redox Signal 11:227–240.
Markowicz-Piasecka M, Sikora J, Szyd?owska A, Skupien A, Mikiciuk-Olasik E, and Huttunen KM (2017) Metformin - a future therapy for neurodegenerative diseases: theme: drug discovery, development and delivery in Alzheimer’s disease: guest editor: Davide Brambilla. Pharm Res 34:2614–2627.
Marrone G, Maeso-Díaz R, García-Cardena G, Abraldes JG, García-Pagán JC, Bosch J, and Gracia-Sancho J (2015) KLF2 exerts antifibrotic and vasoprotective effects in cirrhotic rat livers: behind the molecular mechanisms of statins. Gut 64: 1434–1443.
Martín-de-Saavedra MD, Budni J, Cunha MP, Gómez-Rangel V, Lorrio S, Del Barrio L, Lastres-Becker I, Parada E, Tordera RM, Rodrigues AL, et al. (2013) Nrf2 participates in depressive disorders through an anti-inflammatory mechanism. Psychoneuroendocrinology 38:2010–2022.
Marzec JM, Christie JD, Reddy SP, Jedlicka AE, Vuong H, Lanken PN, Aplenc R, Yamamoto T, Yamamoto M, Cho HY, et al. (2007) Functional polymorphisms in the transcription factor NRF2 in humans increase the risk of acute lung injury. FASEB J 21:2237–2246.
McCubrey JA, Steelman LS, Bertrand FE, Davis NM, Sokolosky M, Abrams SL, Montalto G, D’Assoro AB, Libra M, Nicoletti F, et al. (2014) GSK-3 as potential target for therapeutic intervention in cancer. Oncotarget 5:2881–2911.
McGuire VA, Ruiz-Zorrilla Diez T, Emmerich CH, Strickson S, Ritorto MS, Sutavani RV, Weib A, Houslay KF, Knebel A, Meakin PJ, et al. (2016) Dimethyl fumarate blocks pro-inflammatory cytokine production via inhibition of TLR induced M1 and K63 ubiquitin chain formation. Sci Rep 6:31159.
Meakin PJ, Chowdhry S, Sharma RS, Ashford FB, Walsh SV, McCrimmon RJ, Dinkova-Kostova AT, Dillon JF, Hayes JD, and Ashford ML (2014) Susceptibility of Nrf2-null mice to steatohepatitis and cirrhosis upon consumption of a high-fat diet is associated with oxidative stress, perturbation of the unfolded protein response, and disturbance in the expression of metabolic enzymes but not with insulin resistance. Mol Cell Biol 34:3305–3320.
Meissner M, Doll M, Hrgovic I, Reichenbach G, König V, Hailemariam-Jahn T, Gille J, and Kaufmann R (2011) Suppression of VEGFR2 expression in human endothelial cells by dimethylfumarate treatment: evidence for anti-angiogenic action. J Invest Dermatol 131:1356–1364.
Menche J, Sharma A, Kitsak M, Ghiassian SD, Vidal M, Loscalzo J, and Barabási AL (2015) Disease networks: uncovering disease-disease relationships through the incomplete interactome. Science 347:1257601.
Milkovic L, Zarkovic N, and Saso L (2017) Controversy about pharmacological modulation of Nrf2 for cancer therapy. Redox Biol 12:727–732.
Mitsuishi Y, Taguchi K, Kawatani Y, Shibata T, Nukiwa T, Aburatani H, Yamamoto M, and Motohashi H (2012) Nrf2 redirects glucose and glutamine into anabolic pathways in metabolic reprogramming. Cancer Cell 22:66–79.
Moon SJ, Park JS, Woo YJ, Lim MA, Kim SM, Lee SY, Kim EK, Lee HJ, Lee WS, Park SH, et al. (2014) Rebamipide suppresses collagen-induced arthritis through reciprocal regulation of th17/treg cell differentiation and heme oxygenase 1 induction. Arthritis Rheumatol 66:874–885.
Morgan MJ and Liu ZG (2011) Crosstalk of reactive oxygen species and NF-kB signaling. Cell Res 21:103–115.
Morgan PE, Sturgess AD, Hennessy A, and Davies MJ (2007) Serum protein oxidation and apolipoprotein CIII levels in people with systemic lupus erythematosus with and without nephritis. Free Radic Res 41:1301–1312.
Mrowietz U, Christophers E, and Altmeyer P (1998) Treatment of psoriasis with fumaric acid esters: results of a prospective multicentre study: German Multicentre study. Br J Dermatol 138:456–460.
Mrowietz U, Szepietowski JC, Loewe R, van de Kerkhof P, Lamarca R, Ocker WG, Tebbs VM, and Pau-Charles I (2017) Efficacy and safety of LAS41008 (dimethyl fumarate) in adults with moderate-to-severe chronic plaque psoriasis: a randomized, double-blind, Fumaderm®- and placebo-controlled trial (BRIDGE). Br J Dermatol 176:615–623.
Na LX, Li Y, Pan HZ, Zhou XL, Sun DJ, Meng M, Li XX, and Sun CH (2013) Curcuminoids exert glucose-lowering effect in type 2 diabetes by decreasing serum free fatty acids: a double-blind, placebo-controlled trial. Mol Nutr Food Res 57: 1569–1577.
Napetschnig J and Wu H (2013) Molecular basis of NF-kB signaling. Annu Rev Biophys 42:443–468.
Nasiri HR, Linge S, and Ullmann D (2016) Thermodynamic profiling of inhibitors of Nrf2:Keap1 interactions. Bioorg Med Chem Lett 26:526–529.
Natarajan VT, Singh A, Kumar AA, Sharma P, Kar HK, Marrot L, Meunier JR, Natarajan K, Rani R, and Gokhale RS (2010) Transcriptional upregulation of Nrf2- dependent phase II detoxification genes in the involved epidermis of vitiligo vulgaris. J Invest Dermatol 130:2781–2789.
Nef HM, Möllmann H, Troidl C, Kostin S, Böttger T, Voss S, Hilpert P, Krause N, Weber M, Rolf A, et al. (2008) Expression profiling of cardiac genes in Tako-Tsubo cardiomyopathy: insight into a new cardiac entity. J Mol Cell Cardiol 44:395–404.
Nesti L and Natali A (2017) Metformin effects on the heart and the cardiovascular system: a review of experimental and clinical data. Nutr Metab Cardiovasc Dis 27: 657–669.
Ockenfels HM, Schultewolter T, Ockenfels G, Funk R, and Goos M (1998) The antipsoriatic agent dimethylfumarate immunomodulates T-cell cytokine secretion and inhibits cytokines of the psoriatic cytokine network. Br J Dermatol 139: 390–395.
Okano Y, Nezu U, Enokida Y, Lee MT, Kinoshita H, Lezhava A, Hayashizaki Y, Morita S, Taguri M, Ichikawa Y, et al. (2013) SNP (-617C.A) in ARE-like loci of the NRF2 gene: a new biomarker for prognosis of lung adenocarcinoma in Japanese non-smoking women. PLoS One 8:e73794.
Olayanju A, Copple IM, Bryan HK, Edge GT, Sison RL, Wong MW, Lai ZQ, Lin ZX, Dunn K, Sanderson CM, et al. (2015) Brusatol provokes a rapid and transient inhibition of Nrf2 signaling and sensitizes mammalian cells to chemical toxicity: implications for therapeutic targeting of Nrf2. Free Radic Biol Med 78:202–212.
Onderdijk AJ, Balak DM, Baerveldt EM, Florencia EF, Kant M, Laman JD, van IJcken WF, Racz E, de Ridder D, Thio HB, et al. (2014) Regulated genes in psoriatic skin during treatment with fumaric acid esters. Br J Dermatol 171:732–741.
Onyiah JC, Sheikh SZ, Maharshak N, Otterbein LE, and Plevy SE (2014) Heme oxygenase-1 and carbon monoxide regulate intestinal homeostasis and mucosal immune responses to the enteric microbiota. Gut Microbes 5:220–224.
Orena S, Owen J, Jin F, Fabian M, Gillitt ND, and Zeisel SH (2015) Extracts of fruits and vegetables activate the antioxidant response element in IMR-32 cells. J Nutr 145:2006–2011.
Owen MR, Doran E, and Halestrap AP (2000) Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem J 348:607–614.
Padmanabhan B, Tong KI, Ohta T, Nakamura Y, Scharlock M, Ohtsuji M, Kang MI, Kobayashi A, Yokoyama S, and Yamamoto M (2006) Structural basis for defects of Keap1 activity provoked by its point mutations in lung cancer. Mol Cell 21: 689–700.
Pajares M, Cuadrado A, and Rojo AI (2017) Modulation of proteostasis by transcription factor NRF2 and impact in neurodegenerative diseases. Redox Biol 11: 543–553.
Pajares M, Jiménez-Moreno N, Dias IH, Debelec B, Vucetic M, Fladmark KE, Basaga H, Ribaric S, Milisav I, and Cuadrado A (2015) Redox control of protein degradation. Redox Biol 6:409–420.
Pajares M, Jiménez-Moreno N, García-Yagüe AJ, Escoll M, de Ceballos ML, Van Leuven F, Rábano A, Yamamoto M, Rojo AI, and Cuadrado A (2016) Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 12: 1902–1916.
Palomo V and Martinez A (2017) Glycogen synthase kinase 3 (GSK-3) inhibitors: a patent update (2014-2015). Expert Opin Ther Pat 27:657–666.
Panieri E and Santoro MM (2016) ROS homeostasis and metabolism: a dangerous liason in cancer cells. Cell Death Dis 7:e2253.
Pantan R, Tocharus J, Suksamrarn A, and Tocharus C (2016) Synergistic effect of atorvastatin and cyanidin-3-glucoside on angiotensin II-induced inflammation in vascular smooth muscle cells. Exp Cell Res 342:104–112.
Pareek TK, Belkadi A, Kesavapany S, Zaremba A, Loh SL, Bai L, Cohen ML, Meyer C, Liby KT, Miller RH, et al. (2011) Triterpenoid modulation of IL-17 and Nrf-2 expression ameliorates neuroinflammation and promotes remyelination in autoimmune encephalomyelitis. Sci Rep 1:201.
Park SY, Lee SW, Shin HK, Chung WT, Lee WS, Rhim BY, Hong KW, and Kim CD (2010) Cilostazol enhances apoptosis of synovial cells from rheumatoid arthritis patients with inhibition of cytokine formation via Nrf2-linked heme oxygenase 1 induction. Arthritis Rheum 62:732–741.
Pedruzzi LM, Cardozo LF, Daleprane JB, Stockler-Pinto MB, Monteiro EB, Leite M Jr, Vaziri ND, and Mafra D (2015) Systemic inflammation and oxidative stress in hemodialysis patients are associated with down-regulation of Nrf2. J Nephrol 28: 495–501.
Peng H, Guerau-de-Arellano M, Mehta VB, Yang Y, Huss DJ, Papenfuss TL, LovettRacke AE, and Racke MK (2012) Dimethyl fumarate inhibits dendritic cell maturation via nuclear factor kB (NF-kB) and extracellular signal-regulated kinase 1 and 2 (ERK1/2) and mitogen stress-activated kinase 1 (MSK1) signaling. J Biol Chem 287:28017–28026.
Pergola PE, Raskin P, Toto RD, Meyer CJ, Huff JW, Grossman EB, Krauth M, Ruiz S, Audhya P, Christ-Schmidt H, et al.; BEAM Study Investigators (2011) Bardoxolone methyl and kidney function in CKD with type 2 diabetes. N Engl J Med 365:327–336.
Pilar Valdecantos M, Prieto-Hontoria PL, Pardo V, Módol T, Santamaría B, Weber M, Herrero L, Serra D, Muntané J, Cuadrado A, et al. (2015) Essential role of Nrf2 in the protective effect of lipoic acid against lipoapoptosis in hepatocytes. Free Radic Biol Med 84:263–278.
Piñero J, Bravo À, Queralt-Rosinach N, Gutiérrez-Sacristán A, Deu-Pons J, Centeno E, García-García J, Sanz F, and Furlong LI (2017) DisGeNET: a comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res 45:D833–D839.
Pitha-Rowe I, Liby K, Royce D, and Sporn M (2009) Synthetic triterpenoids attenuate cytotoxic retinal injury: cross-talk between Nrf2 and PI3K/AKT signaling through inhibition of the lipid phosphatase PTEN. Invest Ophthalmol Vis Sci 50: 5339–5347.
Qiu P, Man S, Li J, Liu J, Zhang L, Yu P, and Gao W (2016) Overdose intake of curcumin initiates the unbalanced state of bodies. J Agric Food Chem 64: 2765–2771.
Rachakonda G, Xiong Y, Sekhar KR, Stamer SL, Liebler DC, and Freeman ML (2008) Covalent modification at Cys151 dissociates the electrophile sensor Keap1 from the ubiquitin ligase CUL3. Chem Res Toxicol 21:705–710.
Rada P, Rojo AI, Chowdhry S, McMahon M, Hayes JD, and Cuadrado A (2011) SCF/ beta-TrCP promotes glycogen synthase kinase 3-dependent degradation of the Nrf2 transcription factor in a Keap1-independent manner. Mol Cell Biol 31:1121–1133.
Rada P, Rojo AI, Evrard-Todeschi N, Innamorato NG, Cotte A, Jaworski T, TobónVelasco JC, Devijver H, García-Mayoral MF, Van Leuven F, et al. (2012) Structural and functional characterization of Nrf2 degradation by the glycogen synthase kinase 3/b-TrCP axis. Mol Cell Biol 32:3486–3499.
Ramadori P, Drescher H, Erschfeld S, Schumacher F, Berger C, Fragoulis A, Schenkel J, Kensler TW, Wruck CJ, Trautwein C, et al. (2016) Hepatocyte-specific Keap1 deletion reduces liver steatosis but not inflammation during non-alcoholic steatohepatitis development. Free Radic Biol Med 91:114–126.
Ramsey CP, Glass CA, Montgomery MB, Lindl KA, Ritson GP, Chia LA, Hamilton RL, Chu CT, and Jordan-Sciutto KL (2007) Expression of Nrf2 in neurodegenerative diseases. J Neuropathol Exp Neurol 66:75–85.
Rangasamy T, Cho CY, Thimmulappa RK, Zhen L, Srisuma SS, Kensler TW, Yamamoto M, Petrache I, Tuder RM, and Biswal S (2004) Genetic ablation of Nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J Clin Invest 114:1248–1259.
Ranjan N, Fulcrand G, King A, Brown J, Jiang X, Leng F, and Arya DP (2014) Selective inhibition of bacterial topoisomerase I by alkynyl-bisbenzimidazoles. MedChemComm 5:816–825.
Reccia I, Kumar J, Akladios C, Virdis F, Pai M, Habib N, and Spalding D (2017) Nonalcoholic fatty liver disease: a sign of systemic disease. Metabolism 72:94–108.
Ren D, Villeneuve NF, Jiang T, Wu T, Lau A, Toppin HA, and Zhang DD (2011) Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proc Natl Acad Sci USA 108:1433–1438.
Rena G, Hardie DG, and Pearson ER (2017) The mechanisms of action of metformin. Diabetologia 60:1577–1585.
Reuter S, Gupta SC, Chaturvedi MM, and Aggarwal BB (2010) Oxidative stress, inflammation, and cancer: how are they linked? Free Radic Biol Med 49:1603–1616.
Richardson BG, Jain AD, Speltz TE, and Moore TW (2015) Non-electrophilic modulators of the canonical Keap1/Nrf2 pathway. Bioorg Med Chem Lett 25:2261–2268.
Roberts CA, Dickinson AK, and Taams LS (2015) The interplay between monocytes/ macrophages and CD4(+) T cell subsets in rheumatoid arthritis. Front Immunol 6: 571.
Rojo AI, Innamorato NG, Martín-Moreno AM, De Ceballos ML, Yamamoto M, and Cuadrado A (2010) Nrf2 regulates microglial dynamics and neuroinflammation in experimental Parkinson’s disease. Glia 58:588–598.
Rojo AI, McBean G, Cindric M, Egea J, López MG, Rada P, Zarkovic N, and Cuadrado A (2014a) Redox control of microglial function: molecular mechanisms and functional significance. Antioxid Redox Signal 21:1766–1801.
Rojo AI, Rada P, Mendiola M, Ortega-Molina A, Wojdyla K, Rogowska-Wrzesinska A, Hardisson D, Serrano M, and Cuadrado A (2014b) The PTEN/NRF2 axis promotes human carcinogenesis. Antioxid Redox Signal 21:2498–2514.
Rushworth SA, Zaitseva L, Murray MY, Shah NM, Bowles KM, and MacEwan DJ (2012) The high Nrf2 expression in human acute myeloid leukemia is driven by NF-kB and underlies its chemo-resistance. Blood 120:5188–5198.
Ryoo IG, Lee SH, and Kwak MK (2016) Redox modulating NRF2: a potential mediator of cancer stem cell resistance. Oxid Med Cell Longev 2016:2428153.
Saddawi-Konefka R, Seelige R, Gross ET, Levy E, Searles SC, Washington A Jr, Santosa EK, Liu B, O’Sullivan TE, Harismendy O, et al. (2016) Nrf2 induces IL-17D to mediate tumor and virus surveillance. Cell Reports 16:2348–2358.
Saito R, Suzuki T, Hiramoto K, Asami S, Naganuma E, Suda H, Iso T, Yamamoto H, Morita M, Baird L, et al. (2015) Characterizations of three major cysteine sensors of Keap1 in stress response. Mol Cell Biol 36:271–284.
Saracino AM and Orteu CH (2017) Severe recalcitrant cutaneous manifestations in systemic lupus erythematosus successfully treated with fumaric acid esters. Br J Dermatol 176:472–480.
Sato M, Aoki T, Inoue H, Tanaka T, and Kunishima N (2013) Keap1 Protein Binding Compound, Cristal of Complex Between the Same and Keap1 Protein, and Method for Producing the Same, Toray Industries, Tokyo. Sato N, Takasaka N, Yoshida M, Tsubouchi K, Minagawa S, Araya J, Saito N, Fujita Y, Kurita Y, Kobayashi K, et al. (2016) Metformin attenuates lung fibrosis development via NOX4 suppression. Respir Res 17:107.
Satoh T, McKercher SR, and Lipton SA (2013) Nrf2/ARE-mediated antioxidant actions of pro-electrophilic drugs. Free Radic Biol Med 65:645–657.
Scannevin RH, Chollate S, Jung MY, Shackett M, Patel H, Bista P, Zeng W, Ryan S, Yamamoto M, Lukashev M, et al. (2012) Fumarates promote cytoprotection of central nervous system cells against oxidative stress via the nuclear factor (erythroid-derived 2)-like 2 pathway. J Pharmacol Exp Ther 341:274–284.
Schilling S, Goelz S, Linker R, Luehder F, and Gold R (2006) Fumaric acid esters are effective in chronic experimental autoimmune encephalomyelitis and suppress macrophage infiltration. Clin Exp Immunol 145:101–107.
Schimrigk S, Brune N, Hellwig K, Lukas C, Bellenberg B, Rieks M, Hoffmann V, Pohlau D, and Przuntek H (2006) Oral fumaric acid esters for the treatment of active multiple sclerosis: an open-label, baseline-controlled pilot study. Eur J Neurol 13:604–610.
Schipper HM, Song W, Zukor H, Hascalovici JR, and Zeligman D (2009) Heme oxygenase-1 and neurodegeneration: expanding frontiers of engagement. J Neurochem 110:469–485.
Schuiveling M, Vazirpanah N, Radstake TRDJ, Zimmermann M, and Broen JCA (2017) Metformin, a new era for an old drug in the treatment of immune mediated disease? Curr Drug Targets [published ahead of print]. Schulze-Topphoff U, Varrin-Doyer M, Pekarek K, Spencer CM, Shetty A, Sagan SA, Cree BA, Sobel RA, Wipke BT, Steinman L, et al. (2016) Dimethyl fumarate treatment induces adaptive and innate immune modulation independent of Nrf2. Proc Natl Acad Sci USA 113:4777–4782.
Schumacker PT (2006) Reactive oxygen species in cancer cells: live by the sword, die by the sword. Cancer Cell 10:175–176.
Sha LK, Sha W, Kuchler L, Daiber A, Giegerich AK, Weigert A, Knape T, Snodgrass R, Schröder K, Brandes RP, et al. (2015) Loss of Nrf2 in bone marrow-derived macrophages impairs antigen-driven CD8(+) T cell function by limiting GSH and Cys availability. Free Radic Biol Med 83:77–88.
Shanmugam G, Narasimhan M, Conley RL, Sairam T, Kumar A, Mason RP, Sankaran R, Hoidal JR, and Rajasekaran NS (2017a) Chronic endurance exercise impairs cardiac structure and function in middle-aged mice with impaired Nrf2 signaling. Front Physiol 8:268.
Shanmugam G, Narasimhan M, Tamowski S, Darley-Usmar V, and Rajasekaran NS (2017b) Constitutive activation of Nrf2 induces a stable reductive state in the mouse myocardium. Redox Biol 12:937–945.
Sharma A, Rizky L, Stefanovic N, Tate M, Ritchie RH, Ward KW, and de Haan JB (2017) The nuclear factor (erythroid-derived 2)-like 2 (Nrf2) activator dh404 protects against diabetes-induced endothelial dysfunction. Cardiovasc Diabetol 16:33.
Shibata T, Kokubu A, Saito S, Narisawa-Saito M, Sasaki H, Aoyagi K, Yoshimatsu Y, Tachimori Y, Kushima R, Kiyono T, et al. (2011) NRF2 mutation confers malignant potential and resistance to chemoradiation therapy in advanced esophageal squamous cancer. Neoplasia 13:864–873.
Shimizu T, Inoue K, Hachiya H, Shibuya N, Aoki T, and Kubota K (2016) Accumulation of phosphorylated p62 is associated with NF-E2-related factor 2 activation in hepatocellular carcinoma. J Hepatobiliary Pancreat Sci 23:467–471.
Shimoyama Y, Mitsuda Y, Tsuruta Y, Hamajima N, and Niwa T (2014) Polymorphism of Nrf2, an antioxidative gene, is associated with blood pressure and cardiovascular mortality in hemodialysis patients. Int J Med Sci 11:726–731.
Shimozono R, Asaoka Y, Yoshizawa Y, Aoki T, Noda H, Yamada M, Kaino M, and Mochizuki H (2013) Nrf2 activators attenuate the progression of nonalcoholic steatohepatitis-related fibrosis in a dietary rat model. Mol Pharmacol 84:62–70.
Signorelli SS, Li Volsi G, Fiore V, Mangiafico M, Barbagallo I, Parenti R, Rizzo M, and Li Volti G (2016) Plasma heme oxygenase-1 is decreased in peripheral artery disease patients. Mol Med Rep 14:3459–3463.
Silva T, Reis J, Teixeira J, and Borges F (2014) Alzheimer’s disease, enzyme targets and drug discovery struggles: from natural products to drug prototypes. Ageing Res Rev 15:116–145.
Singh A, Misra V, Thimmulappa RK, Lee H, Ames S, Hoque MO, Herman JG, Baylin SB, Sidransky D, Gabrielson E, et al. (2006) Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med 3:e420.
Singh A, Venkannagari S, Oh KH, Zhang YQ, Rohde JM, Liu L, Nimmagadda S, Sudini K, Brimacombe KR, Gajghate S, et al. (2016) Small molecule inhibitor of NRF2 selectively intervenes therapeutic resistance in KEAP1-deficient NSCLC tumors. ACS Chem Biol 11:3214–3225.
Siomek A (2012) NF-kB signaling pathway and free radical impact. Acta Biochim Pol 59:323–331.
Song P, Li K, Liu L, Wang X, Jian Z, Zhang W, Wang G, Li C, and Gao T (2016) Genetic polymorphism of the Nrf2 promoter region is associated with vitiligo risk in Han Chinese populations. J Cell Mol Med 20:1840–1850.
Stelzer G, Rosen N, Plaschkes I, Zimmerman S, Twik M, Fishilevich S, Stein TI, Nudel R, Lieder I, Mazor Y, et al. (2016) The GeneCards suite: from gene data mining to disease genome sequence analyses. Curr Protoc Bioinformatics 54: 1.30.1–1.30.33.
Stocker R, Yamamoto Y, McDonagh AF, Glazer AN, and Ames BN (1987) Bilirubin is an antioxidant of possible physiological importance. Science 235:1043–1046.
Strom J and Chen QM (2017) Loss of Nrf2 promotes rapid progression to heart failure following myocardial infarction. Toxicol Appl Pharmacol 327:52–58.
Sun H, Zhu J, Lin H, Gu K, and Feng F (2017) Recent progress in the development of small molecule Nrf2 modulators: a patent review (2012-2016). Expert Opin Ther Pat 27:763–785.
Taguchi K and Yamamoto M (2017) The KEAP1-NRF2 system in cancer. Front Oncol 7:85.
Tahvili S, Zandieh B, and Amirghofran Z (2015) The effect of dimethyl fumarate on gene expression and the level of cytokines related to different T helper cell subsets in peripheral blood mononuclear cells of patients with psoriasis. Int J Dermatol 54: e254–e260.
Takahashi Y, Kobayashi Y, Kawata K, Kawamura K, Sumiyoshi S, Noritake H, Watanabe S, Chida T, Souda K, Sakaguchi T, et al. (2014) Does hepatic oxidative stress enhance activation of nuclear factor-E2-related factor in patients with nonalcoholic steatohepatitis? Antioxid Redox Signal 20:538–543.
Tan SM, Sharma A, Stefanovic N, Yuen DY, Karagiannis TC, Meyer C, Ward KW, Cooper ME, and de Haan JB (2014) Derivative of bardoxolone methyl, dh404, in an inverse dose-dependent manner lessens diabetes-associated atherosclerosis and improves diabetic kidney disease. Diabetes 63:3091–3103.
Tang X, Lin CC, Spasojevic I, Iversen ES, Chi JT, and Marks JR (2014) A joint analysis of metabolomics and genetics of breast cancer. Breast Cancer Res 16:415.
Tarumoto T, Nagai T, Ohmine K, Miyoshi T, Nakamura M, Kondo T, Mitsugi K, Nakano S, Muroi K, Komatsu N, et al. (2004) Ascorbic acid restores sensitivity to imatinib via suppression of Nrf2-dependent gene expression in the imatinibresistant cell line. Exp Hematol 32:375–381.
Thimmulappa RK, Lee H, Rangasamy T, Reddy SP, Yamamoto M, Kensler TW, and Biswal S (2006a) Nrf2 is a critical regulator of the innate immune response and survival during experimental sepsis. J Clin Invest 116:984–995.
Thimmulappa RK, Scollick C, Traore K, Yates M, Trush MA, Liby KT, Sporn MB, Yamamoto M, Kensler TW, and Biswal S (2006b) Nrf2-dependent protection from LPS induced inflammatory response and mortality by CDDO-Imidazolide. Biochem Biophys Res Commun 351:883–889.
Thomson L, Tenopoulou M, Lightfoot R, Tsika E, Parastatidis I, Martinez M, Greco TM, Doulias PT, Wu Y, Tang WH, et al. (2012) Immunoglobulins against tyrosinenitrated epitopes in coronary artery disease. Circulation 126:2392–2401.
Todorovic M, Newman JR, Shan J, Bentley S, Wood SA, Silburn PA, and Mellick GD (2015) Comprehensive assessment of genetic sequence variants in the antioxidant ‘master regulator’ NRF2 in idiopathic Parkinson’s disease. PLoS One 10:e0128030.
Tong KI, Padmanabhan B, Kobayashi A, Shang C, Hirotsu Y, Yokoyama S, and Yamamoto M (2007) Different electrostatic potentials define ETGE and DLG motifs as hinge and latch in oxidative stress response. Mol Cell Biol 27:7511–7521.
Tsai HH, Lai HY, Chen YC, Li CF, Huang HS, Liu HS, Tsai YS, and Wang JM (2017) Metformin promotes apoptosis in hepatocellular carcinoma through the CEBPDinduced autophagy pathway. Oncotarget 8:13832–13845.
Tsuchida K, Tsujita T, Hayashi M, Ojima A, Keleku-Lukwete N, Katsuoka F, Otsuki A, Kikuchi H, Oshima Y, Suzuki M, et al. (2017) Halofuginone enhances the chemosensitivity of cancer cells by suppressing NRF2 accumulation. Free Radic Biol Med 103:236–247.
Tu J, Zhang X, Zhu Y, Dai Y, Li N, Yang F, Zhang Q, Brann DW, and Wang R (2015) Cell-permeable peptide targeting the Nrf2-Keap1 interaction: a potential novel therapy for global cerebral ischemia. J Neurosci 35:14727–14739.
Tung MC, Lin PL, Wang YC, He TY, Lee MC, Yeh SD, Chen CY, and Lee H (2015) Mutant p53 confers chemoresistance in non-small cell lung cancer by upregulating Nrf2. Oncotarget 6:41692–41705.
Türei D, Papp D, Fazekas D, Földvári-Nagy L, Módos D, Lenti K, Csermely P, and Korcsmáros T (2013) NRF2-ome: an integrated web resource to discover protein interaction and regulatory networks of NRF2. Oxid Med Cell Longev 2013: 737591.
Tzima S, Victoratos P, Kranidioti K, Alexiou M, and Kollias G (2009) Myeloid heme oxygenase-1 regulates innate immunity and autoimmunity by modulating IFNbeta production. J Exp Med 206:1167–1179.
Ungvari Z, Bagi Z, Feher A, Recchia FA, Sonntag WE, Pearson K, de Cabo R, and Csiszar A (2010) Resveratrol confers endothelial protection via activation of the antioxidant transcription factor Nrf2. Am J Physiol Heart Circ Physiol 299: H18–H24.
Ungvari Z, Bailey-Downs L, Gautam T, Jimenez R, Losonczy G, Zhang C, Ballabh P, Recchia FA, Wilkerson DC, Sonntag WE, et al. (2011) Adaptive induction of NF-E2- related factor-2-driven antioxidant genes in endothelial cells in response to hyperglycemia. Am J Physiol Heart Circ Physiol 300:H1133–H1140.
Uruno A, Furusawa Y, Yagishita Y, Fukutomi T, Muramatsu H, Negishi T, Sugawara A, Kensler TW, and Yamamoto M (2013) The Keap1-Nrf2 system prevents onset of diabetes mellitus. Mol Cell Biol 33:2996–3010.
Uruno A, Yagishita Y, and Yamamoto M (2015) The Keap1-Nrf2 system and diabetes mellitus. Arch Biochem Biophys 566:76–84.
van Horssen J, Drexhage JA, Flor T, Gerritsen W, van der Valk P, and de Vries HE (2010) Nrf2 and DJ1 are consistently upregulated in inflammatory multiple sclerosis lesions. Free Radic Biol Med 49:1283–1289.
van Muiswinkel FL, de Vos RA, Bol JG, Andringa G, Jansen Steur EN, Ross D, Siegel D, and Drukarch B (2004) Expression of NAD(P)H:quinone oxidoreductase in the normal and Parkinsonian substantia nigra. Neurobiol Aging 25:1253–1262.
Vidal M, Cusick ME, and Barabási AL (2011) Interactome networks and human disease. Cell 144:986–998.
von Otter M, Bergström P, Quattrone A, De Marco EV, Annesi G, Söderkvist P, Wettinger SB, Drozdzik M, Bialecka M, Nissbrandt H, et al. (2014) Genetic associations of Nrf2-encoding NFE2L2 variants with Parkinson’s disease: a multicenter study. BMC Med Genet 15:131.
von Otter M, Landgren S, Nilsson S, Celojevic D, Bergström P, Håkansson A, Nissbrandt H, Drozdzik M, Bialecka M, Kurzawski M, et al. (2010a) Association of Nrf2-encoding NFE2L2 haplotypes with Parkinson’s disease. BMC Med Genet 11: 36.
von Otter M, Landgren S, Nilsson S, Zetterberg M, Celojevic D, Bergström P, Minthon L, Bogdanovic N, Andreasen N, Gustafson DR, et al. (2010b) Nrf2-encoding NFE2L2 haplotypes influence disease progression but not risk in Alzheimer’s disease and age-related cataract. Mech Ageing Dev 131:105–110.
Wang L, Han J, Shan P, You S, Chen X, Jin Y, Wang J, Huang W, Wang Y, and Liang G (2017a) MD2 blockage protects obesity-induced vascular remodeling via activating AMPK/Nrf2. Obesity (Silver Spring) 25:1532–1539.
Wang R, An J, Ji F, Jiao H, Sun H, and Zhou D (2008) Hypermethylation of the Keap1 gene in human lung cancer cell lines and lung cancer tissues. Biochem Biophys Res Commun 373:151–154.
Wang X, Chen H, Liu J, Ouyang Y, Wang D, Bao W, and Liu L (2015) Association between the NF-E2 related factor 2 gene polymorphism and oxidative stress, antioxidative status, and newly-diagnosed type 2 diabetes mellitus in a Chinese population. Int J Mol Sci 16:16483–16496.
Wang X, de Rivero Vaccari JP, Wang H, Diaz P, German R, Marcillo AE, and Keane RW (2012) Activation of the nuclear factor E2-related factor 2/antioxidant response element pathway is neuroprotective after spinal cord injury. J Neurotrauma 29: 936–945.
Wang X, Hausding M, Weng SY, Kim YO, Steven S, Klein T, Daiber A, and Schuppan D (2018) Gliptins suppress inflammatory macrophage activation to mitigate inflammation, fibrosis, oxidative stress, and vascular dysfunction in models of nonalcoholic steatohepatitis and liver fibrosis. Antioxid Redox Signal. 28:87–109.
Wang X, Li M, Cao Y, Wang J, Zhang H, Zhou X, Li Q, and Wang L (2017b) Tenuigenin inhibits LPS-induced inflammatory responses in microglia via activating the Nrf2-mediated HO-1 signaling pathway. Eur J Pharmacol 809:196–202.
Wang XJ, Hayes JD, Henderson CJ, and Wolf CR (2007) Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha. Proc Natl Acad Sci USA 104:19589–19594.
Wang Y, Huang Y, Xu Y, Ruan W, Wang H, Zhang Y, Saavedra JM, Zhang L, Huang Z, and Pang T (2017c) A dual AMPK/Nrf2 activator reduces brain inflammation after stroke by enhancing microglia M2 polarization. Antioxid Redox Signal 28: 141–163.
Wasik U, Milkiewicz M, Kempinska-Podhorodecka A, and Milkiewicz P (2017) Protection against oxidative stress mediated by the Nrf2/Keap1 axis is impaired in primary biliary cholangitis. Sci Rep 7:44769.
Wen X, Thorne G, Hu L, Joy MS, and Aleksunes LM (2015) Activation of NRF2 signaling in HEK293 cells by a first-in-class direct KEAP1-NRF2 inhibitor. J Biochem Mol Toxicol 29:261–266.
Wenzel P, Kossmann S, Münzel T, and Daiber A (2017) Redox regulation of cardiovascular inflammation: immunomodulatory function of mitochondrial and Noxderived reactive oxygen and nitrogen species. Free Radic Biol Med 109:48–60.
Wenzel P, Oelze M, Coldewey M, Hortmann M, Seeling A, Hink U, Mollnau H, Stalleicken D, Weiner H, Lehmann J, et al. (2007) Heme oxygenase-1: a novel key player in the development of tolerance in response to organic nitrates. Arterioscler Thromb Vasc Biol 27:1729–1735.
Wenzel P, Rossmann H, Müller C, Kossmann S, Oelze M, Schulz A, Arnold N, Simsek C, Lagrange J, Klemz R, et al. (2015) Heme oxygenase-1 suppresses a proinflammatory phenotype in monocytes and determines endothelial function and arterial hypertension in mice and humans. Eur Heart J 36:3437–3446.
Winkel AF, Engel CK, Margerie D, Kannt A, Szillat H, Glombik H, Kallus C, Ruf S, Güssregen S, Riedel J, et al. (2015) Characterization of RA839, a noncovalent small molecule binder to Keap1 and selective activator of Nrf2 signaling. J Biol Chem 290:28446–28455.
Winston JT, Strack P, Beer-Romero P, Chu CY, Elledge SJ, and Harper JW (1999) The SCFbeta-TRCP-ubiquitin ligase complex associates specifically with phosphorylated destruction motifs in IkappaBalpha and beta-catenin and stimulates IkappaBalpha ubiquitination in vitro. Genes Dev 13:270–283.
Wu Q, Wang Q, Mao G, Dowling CA, Lundy SK, and Mao-Draayer Y (2017) Dimethyl fumarate selectively reduces memory T cells and shifts the balance between Th1/ Th17 and Th2 in multiple sclerosis patients. J Immunol 198:3069–3080.
Wu W, Zhao L, Yang P, Zhou W, Li B, Moorhead JF, Varghese Z, Ruan XZ, and Chen Y (2016a) Inflammatory stress sensitizes the liver to atorvastatin-induced injury in ApoE-/- mice. PLoS One 11:e0159512.
Wu WJ, Jia WW, Liu XH, Pan LL, Zhang QY, Yang D, Shen XY, Liu L, and Zhu YZ (2016b) S-propargyl-cysteine attenuates inflammatory response in rheumatoid arthritis by modulating the Nrf2-ARE signaling pathway. Redox Biol 10:157–167.
Xia N, Daiber A, Förstermann U, and Li H (2017) Antioxidant effects of resveratrol in the cardiovascular system. Br J Pharmacol 174:1633–1646.
Xiaobo C, Majidi M, Feng M, Shao R, Wang J, Zhao Y, Baladandayuthapani V, Song J, Fang B, Ji L, et al. (2016) TUSC2(FUS1)-erlotinib induced vulnerabilities in epidermal growth factor receptor (EGFR) wildtype non-small cell lung cancer (NSCLC) targeted by the repurposed drug auranofin. Sci Rep 6:35741.
Xie H, Zhou F, Liu L, Zhu G, Li Q, Li C, and Gao T (2016) Vitiligo: how do oxidative stress-induced autoantigens trigger autoimmunity? J Dermatol Sci 81:3–9.
Xiong M, Li B, Zhu Q, Wang YX, and Zhang HY (2014) Identification of transcription factors for drug-associated gene modules and biomedical implications. Bioinformatics 30:305–309.
Xu IM, Lai RK, Lin SH, Tse AP, Chiu DK, Koh HY, Law CT, Wong CM, Cai Z, Wong CC, et al. (2016a) Transketolase counteracts oxidative stress to drive cancer development. Proc Natl Acad Sci USA 113:E725–E734.
Xu J, Donepudi AC, Moscovitz JE, and Slitt AL (2013) Keap1-knockdown decreases fasting-induced fatty liver via altered lipid metabolism and decreased fatty acid mobilization from adipose tissue. PLoS One 8:e79841.
Xu X, Sun J, Chang X, Wang J, Luo M, Wintergerst KA, Miao L, and Cai L (2016b) Genetic variants of nuclear factor erythroid-derived 2-like 2 associated with the complications in Han descents with type 2 diabetes mellitus of Northeast China. J Cell Mol Med 20:2078–2088.
Xu Z, Wei Y, Gong J, Cho H, Park JK, Sung ER, Huang H, Wu L, Eberhart C, Handa JT, et al. (2014) NRF2 plays a protective role in diabetic retinopathy in mice. Diabetologia 57:204–213.
Xu Z, Zhang F, Sun F, Gu K, Dong S, and He D (2015) Dimethyl fumarate for multiple sclerosis. Cochrane Database Syst Rev 4:CD011076.
Xue P, Hou Y, Chen Y, Yang B, Fu J, Zheng H, Yarborough K, Woods CG, Liu D, Yamamoto M, et al. (2013) Adipose deficiency of Nrf2 in ob/ob mice results in severe metabolic syndrome. Diabetes 62:845–854.
Yamamoto T, Suzuki T, Kobayashi A, Wakabayashi J, Maher J, Motohashi H, and Yamamoto M (2008) Physiological significance of reactive cysteine residues of Keap1 in determining Nrf2 activity. Mol Cell Biol 28:2758–2770.
Yamamoto T, Yoh K, Kobayashi A, Ishii Y, Kure S, Koyama A, Sakamoto T, Sekizawa K, Motohashi H, and Yamamoto M (2004) Identification of polymorphisms in the promoter region of the human NRF2 gene. Biochem Biophys Res Commun 321: 72–79.
Yang M, Yao Y, Eades G, Zhang Y, and Zhou Q (2011) MiR-28 regulates Nrf2 expression through a Keap1-independent mechanism. Breast Cancer Res Treat 129: 983–991.
Yao W, Zhang JC, Ishima T, Dong C, Yang C, Ren Q, Ma M, Han M, Wu J, Suganuma H, et al. (2016) Role of Keap1-Nrf2 signaling in depression and dietary intake of glucoraphanin confers stress resilience in mice. Sci Rep 6:30659.
Yap JL, Worlikar S, MacKerell AD Jr, Shapiro P, and Fletcher S (2011) Smallmolecule inhibitors of the ERK signaling pathway: towards novel anticancer therapeutics. ChemMedChem 6:38–48.
Yoh K, Hirayama A, Ishizaki K, Yamada A, Takeuchi M, Yamagishi S, Morito N, Nakano T, Ojima M, Shimohata H, et al. (2008) Hyperglycemia induces oxidative and nitrosative stress and increases renal functional impairment in Nrf2-deficient mice. Genes Cells 13:1159–1170.
Yore MM, Kettenbach AN, Sporn MB, Gerber SA, and Liby KT (2011) Proteomic analysis shows synthetic oleanane triterpenoid binds to mTOR. PLoS One 6: e22862.
Yoshizaki Y, Mori T, Ishigami-Yuasa M, Kikuchi E, Takahashi D, Zeniya M, Nomura N, Mori Y, Araki Y, Ando F, et al. (2017) Drug-repositioning screening for Keap1- Nrf2 binding inhibitors using fluorescence correlation spectroscopy. Sci Rep 7: 3945.
Yu M, Li H, Liu Q, Liu F, Tang L, Li C, Yuan Y, Zhan Y, Xu W, Li W, et al. (2011) Nuclear factor p65 interacts with Keap1 to repress the Nrf2-ARE pathway. Cell Signal 23:883–892.
Zehir A, Benayed R, Shah RH, Syed A, Middha S, Kim HR, Srinivasan P, Gao J, Chakravarty D, Devlin SM, et al. (2017) Mutational landscape of metastatic cancer revealed from prospective clinical sequencing of 10,000 patients. Nat Med 23: 703–713.
Zeidan TA, Duncan S, Hencken CP, Wynn TA, and Sanrame CN (2014) Prodrugs of Fumarates and Their Use in Treating Various Diseases, Alkermes Pharma Ireland, Dublin, Ireland. Zhang DD (2013) Bardoxolone brings Nrf2-based therapies to light. Antioxid Redox Signal 19:517–518.
Zhang QY, Chu XY, Jiang LH, Liu MY, Mei ZL, and Zhang HY (2017) Identification of non-electrophilic Nrf2 activators from approved drugs. Molecules 22:1–12.
Zhao C, Zhang Y, Liu H, Li P, Zhang H, and Cheng G (2017) Fortunellin protects against high fructose-induced diabetic heart injury in mice by suppressing inflammation and oxidative stress via AMPK/Nrf-2 pathway regulation. Biochem Biophys Res Commun 490:552–559.
Zhao M, Chen H, Ding Q, Xu X, Yu B, and Huang Z (2016) Nuclear factor erythroid 2-related factor 2 deficiency exacerbates lupus nephritis in B6/lpr mice by regulating Th17 cell function. Sci Rep 6:38619.
Zhou X, Menche J, Barabási AL, and Sharma A (2014) Human symptoms-disease network. Nat Commun 5:4212.
本文由福山生物整理翻译,转载请注明出处。
下一篇:Nrf2与癌症特征